факультет биотехнологии и биотехники
кафедра промышленной биотехнологии
Реферат
По “Биотрансформации”
Тема:
Биотрансформация иммобилизованными ферментами.
| |
Работу выполнил: студент V курсу ФБТ Группы БТ-81 Островной Д.В. |
Киев 2002
Оглавление
1. Общие положения
2. Свойства ферментов
3. Строение ферментов
4. Номенклатура ферментов
5. Классификация ферментов и характеристика некоторых групп
6. Локализация ферментов в клетке
7. Методы выделения и очистки ферментов
8. Иммобилизованные ферменты/p>
9.Применение иммобилизованных ферментов в пищевой промышленности
9.1.Получение глюкозо-фруктозных спиртов
9.2. Получение аминокислот
9.3. Получение L-аспарагиновой кислоты
9.4. Получение L-яблочной кислоты
9.5. Получение без лактозного молока
9.6. Получение сахаров из молочной сыворотки
Литература
1. Общие положения
Ферменты (от лат. fermentum - брожение, закваска), специфические белки, присутствующие во всех живых клетках и играющие роль биологических катализаторов. Через их посредство реализуется генетическая информация и осуществляются все процессы обмена веществ и энергии в живых организмах. Ферменты бывают простыми или сложными белками, в состав которых наряду с белковым компонентом (апоферментом) входит небелковая часть - кофермент. Эффективность действия ферментов определяется значительным снижением энергии активации катализируемой реакции в результате образования промежуточных фермент-субстратных комплексов. Присоединение субстратов происходит в активных центрах, которые обладают сходством только с определенными субстратами, чем достигается высокая специфичность (избирательность) действия ферментов. Одна из особенностей ферментов - способность к направленному и регулируемому действию. За счёт этого контролируется согласованность всех звеньев обмена веществ. Эта способность определяется пространственность структурной молекулы ферментов. Она реализуется через изменение скорости действия ферментов и зависит от концентрации соответствующих субстратов и кофакторов, рH среды, температуры, а также от присутствия специфических активаторов и ингибиторов (например, адениловых нуклеотидов, карбонильных, сульфгидрильных соединений и др.). Некоторые ферменты помимо активных центров имеют дополнительные, т.н. аллостерические регуляторные центры. Биосинтез ферментов находится под контролем генов. Различают конститутивные ферменты, постоянно присутствующие в клетках, и индуцируемые ферменты, биосинтез которых активируется под влиянием соответствующих субстратов. Некоторые функционально взаимосвязанные ферменты образуют в клетке структурно организованные полиферментные комплексы. Многие ферменты и ферментные комплексы прочно связаны с мембранами клетки или её органоидов (митохондрий, лизосом, микросом и т.д.) и участвуют в активном транспорте веществ через мембраны.
Известно более 20000 различных ферментов, из которых многие выделены из живых клеток и получены в индивидуальном состоянии. Первый кристаллический фермент (уреаза) выделен американским биохимиком Д.Самнером в 1926 г. Для ряда ферментов изучена последовательность аминокислот и выяснено расположение полипептидных цепей в трёхмерном пространстве. В лабораторных условиях осуществлен искусственный химический синтез фермента рибонуклеазы. Ферменты используют для количественного определения и получения различных веществ, для модификации молекул нуклеиновых кислот методами генной инженерии, диагностики и лечения ряда заболеваний, а также в ряде технологических процессов, применяемых в лёгкой, пищевой и фармацевтической промышленностях.
2. Свойства ферментов
Будучи белками, ферменты обладают всеми их свойствами. Вместе с тем биокатализаторы характеризуются рядом специфических качеств, тоже вытекающих из их белковой природы. Эти качества отличают ферменты от катализаторов обычного типа. Сюда относятся термолабильность ферментов, зависимость их действия от значения рН среды, специфичность и, наконец, подверженность влиянию активаторов и ингибиторов.
Термолабильность ферментов объясняется тем, что температура, с одной стороны, воздействует на белковую часть фермента, приводя при слишком высоких значениях к денатурации белка и снижению каталитической функции, а с другой стороны, оказывает влияние на скорость реакции образования фермент-субстратного комплекса и на все последующие этапы преобразования субстрата, что ведет к усилению катализа.
Зависимость каталитической активности фермента от температуры выражается типичной кривой. До некоторого значения температуры (в среднем до 5О°С) каталитическая активность растет, причем на каждые 10°С примерно в 2 раза повышается скорость преобразования субстрата. В то же время постепенно возрастает количество инактивированного фермента за счет денатурации его белковой части. При температуре выше 50°С денатурация ферментного белка резко усиливается и, хотя скорость реакций преобразования субстрата продолжает расти, активность фермента, выражающаяся количеством превращенного субстрата, падает.
Детальные исследования роста активности ферментов с повышением температуры, проведенные в последнее время, показали более сложный характер этой зависимости, чем указано выше: во многих случаях она не отвечает правилу удвоения активности на каждые 10°С в основном из-за постепенно нарастающих конформационных изменений в молекуле фермента.
Температура, при которой каталитическая активность фермента максимальна, называется его температурным оптимумом.
Температурный оптимум для различных ферментов неодинаков. В общем для ферментов животного происхождения он лежит между 40 и 50°С, а растительного - между 50 и 60°С. Однако есть ферменты с более высоким температурным оптимумом, например, у папаина (фермент растительного происхождения, ускоряющий гидролиз белка) оптимум находится при 8О°С. В то же время у каталазы (фермент, ускоряющий распад Н2О2 до Н2О и О2) оптимальная температура действия находится между 0 и -10°С, а при более высоких температурах происходит энергичное окисление фермента и его инактивация.
Зависимость активности фермента от значения рН среды была установлена свыше 50 лет назад. Для каждого фермента существует оптимальное значение рН среды, при котором он проявляет максимальную активность. Большинство ферментов имеет максимальную активность в зоне рН поблизости от нейтральной точки. В резко кислой или резко щелочной среде хорошо работают лишь некоторые ферменты.
Переход к большей или меньшей (по сравнению с оптимальной) концентрации водородных ионов сопровождается более или менее равномерным падением активности фермента.
Влияние концентрации водородных ионов на каталитическую активность ферментов состоит в воздействии ее на активный центр. При разных значениях рН в реакционной среде активный центр может быть слабее или сильнее ионизирован, больше или меньше экранирован соседними с ним фрагментами полипептидной цепи белковой части фермента и т.п. Кроме того, рН среды влияет на степень ионизации субстрата, фермент-субстратного комплекса и продуктов реакции, оказывает большое влияние на состояние фермента, определяя соотношение в нем катионных и анионных центров, что сказывается на третичной структуре белковой молекулы. Последнее обстоятельство заслуживает особого внимания, так как определенная третичная структура белка-фермента необходима для образования фермент-субстратного комплекса.
Специфичность - одно из наиболее выдающихся качеств ферментов. Эго свойство их было открыто еще в прошлом столетии, когда было сделано наблюдение, что очень близкие по структуре вещества - пространственные изомеры (a- и b-метилглюкозиды) расщепляются по эфирной связи двумя совершенно разными ферментами.
Таким образом, ферменты могут различать химические соединения, отличающиеся друг от друга очень незначительными деталями строения, такими, например, как пространственное расположение метоксильного радикала и атома водорода при 1-м углеродном атоме молекулы метилглюкозида.
По образному выражению, нередко употребляемому в биохимической литературе, фермент подходит к субстрату, как ключ к замку. Это знаменитое правило было сформулировано Э. Фишером в 1894 г. исходя из того, что специфичность действия фермента предопределяется строгим соответствием геометрической структуры субстрата и активного центра фермента.
В 50-е годы нашего столетия это статическое представление было заменено гипотезой Д. Кошланда об индуцированном соответствии субстрата и фермента. Сущность ее сводится к тому, что пространственное соответствие структуры субстрата и активного центра фермента создается в момент их взаимодействия друг с другом, что может быть выряжено формулой “перчатка - рука”. При этом в субстрате уже деформируются некоторые валентные связи и он, таким образом, подготавливается к дальнейшему каталитическому видоизменению, а в молекуле фермента происходят конформационные перестройки. Гипотеза Кошланда, основанная на допущении гибкости активного центра фермента, удовлетворительно объясняла активирование и ингибирование действия ферментов и регуляцию их активности при воздействии различных факторов. В частности, конформационные перестройки в ферменте в процессе изменения его активности Кошланд сравнивал с колебаниями паутины, когда в нее попала добыча (субстрат), подчеркивая этим крайнюю лабильность структуры фермента в процессе каталитического акта.
В настоящее время гипотеза Кошланда постепенно вытесняется гипотезой топохимического соответствия. Сохраняя основные положения гипотезы взаимоиндуцированной настройки субстрата и фермента, она фиксирует внимание на том, что специфичность действия ферментов объясняется в первую очередь узнаванием той части субстрата, которая не изменяется при катализе. Между этой частью субстрата и субстратным центром фермента возникают многочисленные точечные гидрофобные взаимодействия и водородные связи.
3. Строение ферментов
По строению ферменты могут быть однокомпонентными, простыми белками, и двухкомпонентными, сложными белками. Во втором случае в составе фермента обнаруживается добавочная группа небелковой природы.
В разное время возникли различные наименования белковой части и добавочной группы в двухкомпонентных ферментах. Все они до сих пор употребляются в литературе, например:
Фермент в целом Белковая часть Добавочная группа
Симплекс Ферон (носитель) Агон (активная группа)
Холофермент Апофермент Кофермент
Добавочную группу, прочно связанную, не отделяемую от белковой части, называют простетической группой; в отличие от этого добавочную группу, легко отделяющуюся от апофермента и способную к самостоятельному существованию, обычно именуют коферментом.
Химическая природа важнейших коферментов была выяснена в 30-е годы нашего столетия благодаря трудам О. Варбурга, Р. Куна, П. Каррера и др. Оказалось, что роль коферментов в двухкомпонентных ферментах играют большинство витаминов (Е, К, Q, В1, В2, В6 В12, С, Н и др.) или соединений, построенных с участием витаминов (коэнзим А, НАД+ и т. п.). Кроме того, функцию коферментов выполняют такие соединения, как НS-глутатион, многочисленная группа нуклеотидов и их производных, фосфорные эфиры некоторых моносахаридов и ряд других веществ.
Характерной особенностью двухкомпонентных ферментов является то, что ни белковая часть, ни добавочная группа в отдельности не обладают заметной каталитической активностью. Только их комплекс проявляет ферментативные свойства. При этом белок резко повышает каталитическую активность добавочной группы, присущую ей в свободном состоянии в очень малой степени; добавочная же группа стабилизирует белковую часть и делает ее менее уязвимой к денатурирующим агентам. Таким образом, хотя непосредственным исполнителем каталитической функции является простетическая группа, образующая каталитический центр, ее действие немыслимо без участия полипептидных фрагментов белковой части фермента. Более того, в апоферменте есть участок, характеризующийся специфической структурой, избирательно связывающий кофермент. Это так называемый кофермент связывающий домен; его структура у различных апоферментов, соединяющихся с одним и тем же коферментом, очень сходна. Таковы, например, пространственные структуры нуклеотидсвязывающих доменов ряда дегидрогеназ.
Иначе обстоит дело у однокомпонентных ферментов, не имеющих добавочной группы, которая могла бы входить в непосредственный контакт с преобразуемым соединением. Эту функцию выполняет часть белковой молекулы, называемая каталитическим центром. Предполагают, что каталитический центр однокомпонентного фермента представляет собой уникальное сочетание нескольких аминокислотных остатков, располагающихся в определенной части белковой молекулы.
Чаще всего в каталитических центрах однокомпонентных ферментов встречаются остатки сер, гис, три, арг, цис, асп, глу и тир. Радикалы перечисленных аминокислот выполняют здесь ту же функцию, что и кофермент в составе двухкомпонентного фермента.
Аминокислотные остатки, образующие каталитический центр однокомпонентного фермента, расположены в различных точках единой полипептидной цепи. Поэтому каталитический центр возникает в тот момент, когда белковая молекула приобретает присущую ей третичную структуру. Следовательно, изменение третичной структуры фермента под влиянием тех или иных факторов может привести к деформации каталитического центра и изменению ферментативной активности.
Кроме каталитического центра, образованного сочетанием аминокислотных радикалов или присоединением кофермента, у ферментов различают еще два центра: субстратный и аллостерический.
Под субстратным центром понимают участок молекулы фермента, ответственный за присоединение вещества (субстрата), подвергающегося ферментативному превращению. Часто этот участок называют “якорной площадкой” фермента, где, как судно на якорь, становится субстрат. Во многих случаях прикрепление субстрата к ферменту идет за счет взаимодействия с e-аминогрулпой радикала лиз, расположенного в субстратном центре. Эту же роль может выполнять СООН-группа глу, а также НS-группа цис. Однако работы последних лет показали, что гораздо большее значение здесь имеют силы гидрофобных взаимодействий и водородные связи, возникающие между радикалами аминокислотных остатков субстратного центра фермента и соответствующими группировками в молекуле субстрата.
Понятие о каталитическом и субстратном центре не следует абсолютизировать. В реальных ферментах субстратный центр может совпадать (или перекрываться) с каталитическим центром. Более того, каталитический центр может окончательно формироваться в момент присоединения субстрата. Поэтому часто говорят об активном центре фермента, представляющем сочетание первого и второго. Активный центр у ферментов располагается на две щели при двухъядерной структуре, например у лизоцима и рибонуклеазы, или на дне глубокой впадины, как у химотрипсиногена.
Аллостерический центр представляет собой участок молекулы фермента, в результате присоединения к которому определенного низкомолекулярного (а иногда - и высокомолекулярного) вещества изменяется третичная структура белковой молекулы. Вследствие этого изменяется конфигурация активного центра, сопровождающаяся либо увеличением, либо снижением каталитической активности фермента. Это явление лежит в основе так называемой аллостерической регуляции каталитической активности ферментов.
Значения молекулярных масс ферментов колеблются в широких пределах: от нескольких тысяч до нескольких миллионов. В природе насчитывается несколько десятков ферментов, обладающих сравнительно небольшими молекулами (до 50 тыс.). Однако большинство ферментов представлено белками более высокой молекулярной массы, построенными из субъединиц. Так, каталаза (М=25200) содержит в молекуле шесть протомеров с М=42000 каждый. Молекула фермента, ускоряющего реакцию синтеза рибонуклеиновых кислот (РНК-полимераза, М = 400000), состоит из 6 неравных субъединиц. Полная молекула глутаматдегидрогеназы, ускоряющей процесс окисления глутаминовой кислоты (М=336000), построена из 6 субъединиц с М=56000.
Способы компоновки протомеров в мультимеры разнообразны. Крайне важно, что достроенный из субъединиц фермент проявляет максимальную каталитическую активность именно в виде мультимера: диссоциация на протомеры резко снижает активность фермента. Не все ферменты-мультимеры построены исключительно из каталитически активных протомеров. Наряду с каталитическими в их составе отмечены регуляторные субъединицы, как, например, у аспартат-карбамилтрансферазы.
Среди ферментов-мультимеров безусловно преобладают димеры и тетрамеры (их несколько сотен), в меньшей мере распространены гексамеры и октамеры (несколько десятков) и необыкновенно редко встречаются тримеры и пентамеры.
Молекулы ферментов-мультимеров в ряде случаев составлены из субъединиц двух типов, обозначаемых условно как субъединицы типа А и В. Они сходны друг с другом, но отличаются по некоторым деталям первичной и третичной структур. В зависимости от соотношения протомеров типа А и В в мультимере последний может существовать в виде нескольких изомеров, которые называют изозимами. Так, при четырех субъединицах возможны 5 изозимов:
| I |
II |
III |
IV |
V |
| AAAA |
AAAB |
AABB |
ABBB |
BBBB |
В настоящее время интерес к изозимам резко повысился. Оказалось, что кроме генетически детерминированных изозимов существует большая группа ферментов, обладающая множественными формами, возникающими в результате их посттрансляционной модификации. Множественные формы ферментов и изозимы в частности используются сейчас для диагностики болезней в медицине, прогнозирования продуктивности животных подбора родительских пар при скрещивании для обеспечения максимального гетерозиса в потомстве и т. п.
Значение пространственной организации ферментов особенно ярко выявляется при изучении строения так называемых мультиэнзимов, т.е. ферментов, обладающих способностью ускорять одновременно несколько химических реакций и осуществлять сложные превращения субстрата. Примером может служить мультиэнзим, ускоряющий реакцию окислительного декарбоксилирования пировиноградной кислоты. Этот многоферментный комплекс с М=4500000 состоит из трех видов ферментов. Первый из них (E1) ускоряет реакцию декарбоксилирования пировиноградной кислоты. В состав комплекса входит 12 димерных молекул этого фермента (К=19200). Второй и третий ферменты, катализирующие окислительно-восстановительные процессы при окислении пировиноградной кислоты, сосредоточены внутри мультиэнзимного комплекса. Один из них (Е3) представлен шестью димерными молекулами (М=112 000), другой (Е2) - 24 протомерами (М=70000).
В тех случаях, когда мультиэнзимный комплекс обслуживает единый, многоступенчатый процесс биохимических превращений, его называют метаболоном (от слова метаболизм - обмен веществ). Таковы метаболоны гликолиза, биосинтеза ряда аминокислот, цикла дикарбоновых и трикарбоновых кислот и др.
В результате слаженного во времени и пространстве действия всех трех видов входящих в его состав ферментов мультиэнзим с огромной скоростью осуществляет превращение пировиноградной кислоты. Именно в кооперативном характере каталитического процесса и кроется главное отличие биокатализаторов от катализаторов неорганической природы, именно поэтому интенсивность биокатализа в десятки, сотни и тысячи раз превосходит мощность действия неорганических катализаторов.
Сравнительно недавно выявлена еще одна своеобразная черта в строении ферментов: некоторые из них являются полифункциональными, т.е. обладают несколькими энзиматическими активностями, но всего лишь одной полипептидной цепью. Дело в том, что эта единая цепь при формировании третичной структуры образует несколько функционально и стерически обособленных глобулярных участков - доменов, каждый из которых характеризуется своей каталитической активностью.
При изучении мультиэнзимных комплексов и полифункциональных ферментов удалось понять наиболее важную особенность ферментативного катализа, а именно - эстафетную передачу промежуточных продуктов реакции от одного компонента каталитической системы к другому без их высвобождения.
4. Номенклатура ферментов
Ферментология очень долго не располагала строго научной номенклатурой ферментов. Наименования ферментам давали по случайным признакам (тривиальная номенклатура), по названию субстрата (рациональная), по химическому составу фермента, наконец, по типу катализируемой реакции и характеру субстрата.
Примерами тривиальной номенклатуры могут служить названия таких ферментов, как пепсин (от греч. пепсис - пищеварение), трипсин (от греч. трипсис - разжижаю) и папаин (от названия дынного дерева Carica papaja, из сока которого он выделен). По действию все эти ферменты являются протеолитическими, т. е. ускоряют гидролиз протеинов (белков). Характерное название была дано группе окрашенных внутриклеточных ферментов, ускоряющих окислительно-восстановительные реакции в клетке, - цитохромы (от лат. citos - клетка и chroma - цвет).
Наибольшее распространение получила рациональная номенклатура, согласно которой название фермента составляется из названия субстрата характерного окончания -аза. Она была предложена более столетия тому назад, в 1883 г. Э. Дюкло - учеником Л. Пастера. Так, фермент, ускоряющий реакцию гидролиза крахмала, получил название амилаза (от греч. амилон - крахмал), гидролиза жиров - липаза (от греч. липос - жир), белков (протеинов) - протеаза, мочевины - уреаза (от греч. уреа - мочевина) и т. п.
Когда методами аналитической химии были достигнуты известные успехи в расшифровке химической природы простетических групп, возникла новая номенклатура ферментов. Их стали именовать по названию простетической группы, например, геминфермент (простетическая группа - гем), пиридоксаль-фермент (простетическая группа - пиридоксаль) и т.п.
Затем в названии фермента стали указывать как на характер субстрата, так и на тип катализируемой реакции. К примеру, фермент, отнимающий водород от молекулы янтарной кислоты, называют сукцинатдегидрогеназой, подчеркивая этим одновременно и химическую природу субстрата, и отнятие атомов водорода в процессе ферментативного действия:
- 2Н
НООС ¾ СH2 ¾ СН2 ¾ CООН ¾¾¾¾¾® НООС ¾ СН = СН ¾ СООН
Янтарная кислота Дегидрирование
В 1961 г. Международная комиссия по номенклатуре ферментов представила V Международному биологическому конгрессу проект номенклатуры, построенный на строго научных принципах. Проект был утвержден конгрессом, и новая номенклатура прочно вошла в ферментологию. Согласно этой (Московской) номенклатуре название ферментов составляют из химического названия субстрата и названия той реакции, которая осуществляется ферментом. Если химическая реакция, ускоряемая ферментом, сопровождается переносом группировки атомов от субстрата к акцептору, название фермента включает также химическое наименование акцептора.
Например, пиридоксальфермент, катализируюший реакцию переаминирования между L-аланином и a-кетоглутаровой кислотой, называется L-аланин: 2-оксоглутарат аминотрансфераза. В этом названии отмечены сразу три особенности: 1) субстратом является L-аланин; 2) акцептором служит 2-окcоглутаровая кислота; З) от субстрата к акцептору передается аминогруппа.
Названия ферментов по научной номенклатуре неизмеримо выигрывают в точности, но становятся в ряде случаев гораздо сложнее старых, тривиальных. Так, уреаза (тривиальное название), ускоряющая реакцию гидролиза - мочевины на оксид углерода (IV) и аммиак, по научной номенклатуре именуется карбамид - амидогидролазой:
Н2N ¾ СО >2 + Н2О ¾® 2NН3 + СО2
В этом названии дано точное химическое наименование субстрата и указано, что фермент катализирует реакцию гидролиза амидогруппы. Трегалаза, ускоряющая реакцию гидролиза трегалозы, называется трегалоза-1-глюко-гидролазой.
В связи со значительным усложнением научных названий в новой номенклатуре допускается сохранение наряду с новыми старых тривиальных, рабочих названий ферментов. Международной комиссией был составлен детальный список всех известных в то время ферментов, существенно дополненный в 1972 г. при пересмотре как классификации, так и номенклатуры некоторых ферментов, где рядом с новым научным названием каждого фермента приведено старое, а также указан химизм катализируемой ферментом реакции и в некоторых случаях природа фермента. Таким образом, исключается возможность путаницы в наименовании ферментов. В 1964 г. список включал 874 фермента; в последующее время он был существенно дополнен и возрос до 1770 ферментов в 1972 г. и до 2003 ферментов в 1979 г.
Каждому ферменту в указанном списке присвоен индивидуальный номер (шифр). Например, шифр уреазы выражается цифрами 3.5.1.5. Это означает, что уреаза относится к 3-му классу (первая цифра) ферментов, все представители которого катализируют реакции гидролиза. Вторая цифра (5) говорит о том, что уреаза принадлежит к 5-му подклассу этого класса, куда зачислены все ферменты, ускоряющие гидролиз С - N-связей, не являющихся пептидными. Третья цифра шифра (1) указывает на принадлежность уреазы к подподклассу 5-го подкласса, члены которого ускоряют гидролиз линейных амидов, а последняя цифра (5) - порядковый номер уреазы в этом подподклассе.
Упоминавшаяся ранее лактатдегидрогенеза имеет шифр 1.1.1.27, т. е. относится к 1-му классу ферментов (оксидоредуктазы), к 1-му подклассу (оксидоредуктазы, действующие на СН - ОН-группировки в качестве доноров атомов водорода), к 1-му подподклассу (акцептором атомов водорода служит никотинамидадениндинуклеотид) и занимает 27-е место в перечне ферментов упомянутого подподкласса. Таким образом, шифр абсолютно точно указывает место фермента в общем списке. В настоящее время принято в научных публикациях при первом упоминании фермента указывать в скобках его шифр.
5. Классификация ферментов и характеристика некоторых групп
По первой в истории изучения ферментов классификации их делили на две группы: гидролазы, ускоряющие гидролитические реакции, и десмолазы, ускоряющие реакции негидролитического распада. Затем была сделана попытка разбить ферменты на классы по числу субстратов, участвующих в реакции. В соответствии с этим ферменты классифицировали на три группы. 1. Катализирующие превращения двух субстратов одновременно в обоих направлениях: А+В)С+D. 2. Ускоряющие превращения двух субстратов в прямой реакции и одного в обратной: А+В)С. 3. Обеспечивающие каталитическое видоизменение одного субстрата как в прямой, так и в обратной реакции: А)В.
Одновременно развивалось направление, где в основу классификации ферментов был положен тип реакции, подвергающейся каталитическому воздействию. Наряду с ферментами, ускоряющими реакции гидролиза (гидролазы), были изучены ферменты, участвующие в реакциях переноса атомов и атомных групп (феразы), в изомеризации (изомеразы), расщеплении (лиазы), различных синтезах (синтетазы) и т. д. Это направление в классификации ферментов оказалось наиболее плодотворным, так как объединяло ферменты в группы не по надуманным, формальным признакам, а по типу важнейших биохимических процессов, лежащих в основе жизнедеятельности любого организма. По этому принципу все ферменты делят на 6 классов.
1. Оксидоредуктазы - ускоряют реакции окисления - восстановления. 2. Трансферазы - ускоряют реакции переноса функциональных групп и молекулярных остатков. 3. Гидролазы - ускоряют реакции гидролитического распада. 4. Лиазы - ускоряют негидролитическое отщепление от субстратов определенных групп атомов с образованием двойной связи (или присоединяют группы атомов по двойной связи). 5. Изомеразы - ускоряют пространственные или структурные перестройки в пределах одной молекулы. 6. Лигазы - ускоряют реакции синтеза, сопряженные с распадом богатых энергией связей. Эти классы и положены в основу новой научной классификации ферментов.
К классу оксидоредуктаз относят ферменты, катализирующие реакции окисления - восстановления. Окисление протекает как процесс отнятия атомов Н (электронов) от субстрата, а восстановление - как присоединение атомов Н (электронов) к акцептору.
В класс трансфераз входят ферменты, ускоряющие реакции переноса функциональных групп и молекулярных остатков от одного соединения к другому. Это один из наиболее обширных классов: он насчитывает около 500 индивидуальных ферментов. В зависимости от характера переносимых группировок различают фосфотрансферазы, аминотрансферазы, гликозилтрансферазы, ацилтрансферазы, трансферазы, переносящие одноуглеродные остатки (метилтрансферазы, формилтрансферазы), и др. Например, амидазы ускоряют гидролиз амидов кислот. Из них важную роль в биохимических процессах в организме играют уреаза, аспарагиназа и глутаминаза.
Уреаза была одним из первых белков-ферментов, полученным в кристаллическом состоянии. Это однокомпонентный фермент (М=480000), молекула его глобулярна и состоит из 8 равных субъединиц. Уреаза ускоряет гидролиз мочевины до NН3 и СО2.
Характерные черты действия ферментов класса лигаз (синтетаз) выявлены совсем недавно в связи со значительными успехами в изучении механизма синтеза жиров, белков и углеводов: Оказалось, что старые представления об образовании этих соединений, согласно которым они возникают при обращении реакций гидролиза, не соответствуют действительности. Пути их синтеза принципиально иные.
Главная их особенность - сопряженность синтеза с распадом веществ, способных поставлять энергию для осуществления биосинтетического процесса. Одним из таких природных соединений является АТФ. При отрыве от ее молекулы в присутствии лигаз одного или двух концевых остатков фосфорной кислоты выделяется большое количество энергии, используемой для активирования реагирующих веществ. Лигазы же каталитически ускоряют синтез органических соединений из активированных за счет распада АТФ исходных продуктов. Таким образом, к лигазам относятся ферменты, катализирующие соединение друг с другом двух молекул, сопряженное с гидролизом пирофосфатной связи в молекуле АТФ или иного нуклеозидтрифосфата.
Механизм действия лигаз изучен еще недостаточно, но, несомненно, он весьма сложен. В ряде случаев доказано, что одно из участвующих в основной реакции веществ сначала дает промежуточное соединение с фрагментом распадающейся молекулы АТФ, а вслед за этим указанный промежуточный продукт взаимодействует со вторым партнером основной химической реакции с образованием конечного продукта.
6. Локализация ферментов в клетке
Одним из принципиальных отличий ферментов от катализаторов небиологического происхождения является кооперативный характер их действия. На уровне одиночной молекулы фермента кооперативный принцип реализуется в тонком взаимодействии субстратного, активного и аллостерического центров. Однако гораздо большее значение имеет кооперативное осуществление реакций на уровне ансамблей ферментов. Именно благодаря наличию систем ферментов - в виде мультиэнзимных комплексов или еще более сложных образований - метаболонов, обеспечивающих каталитические превращения всех участников единого метаболического цикла - в клетках с большой скоростью осуществляются многостадийные процессы как распада, так и синтеза органических молекул. Ферментативный катализ в многостадийных реакциях идет без выделения промежуточных продуктов: только возникнув, они тут же подвергаются дальнейшим преобразованиям.
Это возможно лишь потому, что в клеточном содержимом ферменты распределены не хаотически, а строго упорядоченно. С современной точки зрения клетка представляется высокоорганизованной системой, в отдельных частях которой осуществляются строго определенные биохимические процессы. В соответствии с приуроченностью их к определенным субклеточным частицам или отсекам (компартментам) клетки в них локализованы те или иные индивидуальные ферменты, мультиэнзимные комплексы, полифункциональные ферменты или сложнейшие метаболоны.
Разнообразные гидролазы и лиазы сосредоточены преимущественно в лизосомах. Внутри этих сравнительно небольших (несколько нанометров в диаметре) пузырьков, ограниченных мембраной от гиалоплазмы клетки, протекают процессы деструкции различных органических соединений до тех простейших структурных единиц, из которых они построены. Сложные ансамбли окислительно-восстановительных ферментов, такие, например, как цитохромная система, находятся в митохондриях. В этих же субклеточных частицах локализован набор ферментов цикла дикарбоновых и трикарбоновых кислот. Ферменты активирования аминокислот распределены в гиалоплазме, но они же есть и в ядре. В гиалоплазме присутствуют многочисленные метаболоны гликолиза, структурно объединенные с таковыми пентозофосфатного цикла, что обеспечивает взаимопереключение дихотомического и апотомического путей распада углеводов. В то же время ферменты, ускоряющие перенос аминокислотных остатков на растущий конец полипептидной цепи и катализирующие некоторые другие реакции в процессе биосинтеза белка, сосредоточены в рибосомальном аппарате клетки. Нуклеотидилтрансферазы, ускоряющие реакцию переноса нуклеотидных остатков при новообразовании нуклеиновых кислот, локализованы в основном в ядерном аппарате клетки. Таким образом, системы ферментов, сосредоточенные в тех или иных структурах, участвуют в осуществлении отдельных циклов реакций. Будучи тонко координированы друг с другом, эти отдельные циклы реакций обеспечивают жизнедеятельность клеток, органов, тканей и организма в целом.
7. Методы выделения и очистки ферментов
Долгое время вполне обоснованно считали, что все ферменты - тела белковой природы. Однако в начале 80-х годов была неожиданно открыта способность низкомолекулярных рибонуклеиновых кислот ускорять реакцию превращения предшественников РНК в функционально значимый продукт, т. е. возникло представление о полирибонуклеотидной природе некоторых ферментов, названных рибозимами.
Хотя уже осуществлен лабораторный синтез ряда ферментов - рибонуклеазы, лизоцима, ферредоксина и цитохрома с, трудно ожидать, что синтетическое получение ферментов получит широкое распространение в ближайшие десятилетия ввиду его сложности и дороговизны, поэтому единственный реальный в настоящее время способ получения ферментов - это выделение их из биологических объектов.
Выделяют ферменты так же, как и другие белки, хотя есть приемы, применяемые преимущественно для ферментов. Из них можно отметить экстракцию глицерином, в котором сохраняются нативные свойства ферментов, а также метод ацетоновых порошков, состоящий в осаждении и быстром обезвоживании при температуре не выше -10°С тканей или вытяжек из них, содержащих ферменты. К их числу относится также получение ферментов путем адсорбции с последующей элюцией (снятием) с адсорбента. Этот метод был введен в химию ферментов А. Я. Данилевским и дал мощный толчок развитию ферментологии. Сейчас адсорбционный метод выделения и очистки ферментов разработан детально. Наряду с ним широко применяют метод ионообменной хроматографии, метод молекулярных сит, электрофорез и особенно изоэлектрофокусирование. Одна из модификаций адсорбционного метода - афинная хроматография, где адсорбентом служит вещество, с которым фермент взаимодействует избирательно. В результате лишь один этот фермент задерживается на колонке, а все сопутствующие ему выходят с током проявителя. Изменяя характер проявителя, исследуемый фермент элюирует с колонки. Этим методом достигают очистки фермента в несколько тысяч раз, применяя всего лишь одноэтажную (аффинная сорбция - элюция) схему выделения.
Для успешного выделения ферментов из клеточного содержимого необходимо очень тонкое измельчение исходного материала, вплоть до разрушения субклеточных структур: лизосом, митохондрий, ядер и др., которые несут в своем составе многие индивидуальные ферменты. Особое внимание при выделении ферментов уделяют проведению всех операций в условиях, исключающих денатурацию белка, так как она всегда связана с потерей ферментативной активности. Этому способствует проведение операций в присутствии защитных добавок, в частности HS-содержащих соединений (цистеина, глутатиона, меркаптоэтанола, цистеамина, дитиотреитола и др.):
HS ¾ CH2 ¾ СН2 ¾ NН HS¾CH2¾CH(ОН) ¾ СН (ОН) ¾ СH2 ¾ SH
Цистеамин Дитиотреитол
Очень важно поддерживать на всех этапах выделения ферментов низкую температуру, так как некоторые из них даже при -80°С теряют активность.
Для оценки гомогенности ферментного препарата прибегают к обычным методам белковой химии. Переломным моментом в усовершенствовании методов получения высокоочищенных, гомогенных препаратов ферментов было открытие способности их кристаллизоваться, осуществленное впервые в 1906 г. А. Д. Розенфельдом (им была получена в виде кристаллов оксидаза из корней редьки) и приобретшее с 1926 г. широкую известность после работы Д. Самнера по получению кристаллической уреазы из бобов канавалии. Нередко о степени чистоты ферментного препарата судят по его биологической активности; если активность при дальнейшей очистке не возрастает, препарат можно считать гомогенным. Из 2003 включенных в список ферментов более 1500 выделено и в той или иной мере очищено, третья часть их закристаллизована, у нескольких сотен выяснена первичная, а у нескольких десятков - третичная структура.
ИММОБИЛИЗОВАННЫЕ ФЕРМЕНТЫ
Принципиально новые перспективы открылись перед прикладной энзимологией в результате создания нового типа биоорганических «катализаторов, так называемых иммобилизованных ферментов, т. е. ферментов, связанных с носителем. Нельсон и Гриффин Цуже в 1916 г. показали, что инвертаза, если адсорбировать ее на угле или на алюмогеле, сохраняет каталитическую активность. Однако целенаправленная разработка такого рода гетерогенных катализаторов на основе ферментов началась лишь в 50-х гг.
Сам термин «иммобилизованные ферменты» узаконен сравнительно недавно [8]. В принципе понятие «иммобилизация» можно понимать шире, чем просто связывание фермента с водонерастворимым носителем, а именно как любое ограничение степеней свободы ферментных молекул (или их, фрагментов). Этого можно достичь даже путем внутримолекулярной «сшивки» белковых глобул низкомолекулярными бифункциональными реагентами или же присоединением фермента к водорастворимому полимеру. Однако такие препараты обычно не называют иммобилизованными; их скорее относят к ферментам, которые модифицированы соответственно «сшивающими» или полимерными реагентами.
Иммобилизованные и модифицированные ферментные препараты, обладают рядом существенных преимуществ (при использовании в прикладных целях) по сравнению с их «нативными» предшестаенниками.
Во-первых, гетерогенный катализатор легко отделить от реакционной среды, что позволяет: 1) остановить реакцию; 2) использовать катализатор повторно; 3) получать продукт, не загрязненный ферментом. Последнее особенно важно в ряде пищевых или фармацевтических производств.
Во-вторых, гетерогенные катализаторы позволяют проводить ферментный процесс непрерывно (например, в проточных реакторах) и регулировать скорость катализируемой реакции (или выход продукта) скоростью потока.
В-третьих, иммобилизация или модификация позволяют целенаправленно изменять свойства фермента, в том числе его специфичность (особенно в отношении к макромолекулярным субстратам), зависимость активности от рН-среды и, что особенно важно, его стабильность по отношению к различного рода денатурирующим воздействиям среды.
Именно эти три момента лежат в основе научно-технического направления, называемого часто «инженерная энзимология». Текущая задача инженерной энзимологии — это разработка (конструирование) биоорганических катализаторов с заданными свойствами на основе ферментов (в том числе с использованием лолиферментных комплексов или даже клеток, искусственно лишенных способности расти). Говоря о «заданных» свойствах, следует понимать, что они продиктованы потребностями практики; это, например, необходимое время службы катализатора при определенных условиях реакции (что зависит от его термо- и кислотостабильности и т. п.), избирательность (специфичность) действия, производительность (каталитическая активность), иммуногенность, токсичность, геометрическая форма гетерогенного катализатора и его механические свойства и др.
Ниже будут рассмотрены важнейшие методические и идейные аспекты инженерной энзимологии, а также ее первые успехи и перспективы развития.
Методы иммобилизации [6, 14]. Известны по крайней мере три типа подходов, позволяющих связывать ферменты с носителем: адсорбционные методы, методы механического включения (захвата) и методы химичеокого (ковалентного) присоединения.
1. Ферменты можно адсорбировать (в результате физических или ионных взаимодействий) на керамике, стекле, силикагеле, оксидах и гидроокеидах металлов, полисахаридах, органических смолах и других носителях. Для усиления адсорбции ферменты иногда химически модифицируют, вводя в поверхностный слой белковой глобулы дополнительные ионные или гидрофобные группы.
2. Весьма широко применяются методы механического включения (захвата) ферментов в полимерные гели, полупроницаемые полимерные микрокапсулы, полые волокна, жидкие мембраны или липосомы, состоящие из поверхностно-активных компонентов и т. п.
3. Особый интерес представляют химические подходы, позволяющие ковалентно присоединять ферменты (через их функциональные группы, несущественные для катализа, в том числе -NН2, -СООН, -SН, -ОН и др.) как к неорганическим носителям (например, к пористому стеклу, керамике, железу), так и к природным материалам (например, к целлюлозе, хитину, декстранам, агарозе) или к синтетическим полимерам (например, к найлону, полистиролу, полдакриламиду, ионообменным смолам).
Для придания носителю достаточно высокой связующей способности его поверхность приходится иногда «активировать». На рис. 1 приведен один из классических методов иммобилизации белка на полисахаридной матрице. На первой стадии носитель окисляют перйодатом калия до появления альдегидных групп затем фермент соединяют с активированным носителем через азометиновые связи и, наконец, для придания большей устойчивости, связь между белком и носителем восстанавливают боргидридом натрия.
Другой пример - сополимеризационный метод (ряс. 2). На первой стадии в ферментную молекулу вводят двойные связи, способные к сополимеризации; например, фермент ацилируют акрилоилхлоридом; затем акрилоилированный фермент вносят в раствор мономера и проводят сополимеризацию. В результате фермент оказывается химически «вшитым» в полимерную сетку геля.
Выбор носителя и иммобилизационного метода зависит от природы фермента и цели, для которой он должен быть применен. В литературе [6, 24] можно найти полезные рекомендации по этим вопросам.
Геометрические формы носителя могут быть самыми различными; широко используются мелкие гранулы, в том числе пористые шарики, трубочки, волокна, пористые пластины (фильтры), полупроницаемые мембраны и т. 'п. В частности, иммобилизационный метод, основанный на захвате фермента в полиэлектрояитные комплексы [12], позволяет (путем изменения рН или ионной силы раствора) переводить носитель из водорастворимого состояния в всдонерастворимое (осадок) и обратно.
Носитель может оказывать существенное влияние на кинетику действия иммобилизованного фермента. Так, необходимо учитывать следующие возможности: внешних и внутренних диффузионных затруднений; стерических препятствий, особенно в реакциях фермента с макромолекулярными субстратами; распределение субстратов, ингибиторов, ионов водорода и других эффекторов между водным раствором и матрицей (за счет электростатических или гидрофобных взаимодействий, водородных связей).
Некоторые из этих эффектов нежелательны, и с ними приходится бороться [4], подбирая другой носитель или условия иммобилизации. С другой стороны, например, эффекты распределения можно целенаправленно использовать для придания катализатору других свойств. Так, при иммобилизации на анионном полиэлектролите (рис. 3, кривая а) происходит концентрирование ионов гидроксония в зоне отрицательно заряженной матрицы, что вызывает сдвиг локального значения рН, характеризующего микроокружение фермента, в «кислую сторону» по сравнению с раствором. В итоге рН-профиль наблюдаемой каталитической активности оказывается смещенным вправо, т. е. в сторону более высоких значений рН буфера. При иммобилизации на катионном полиэлектролите (рис. 3, кривая б) имеет место прямо противоположная зависимость.
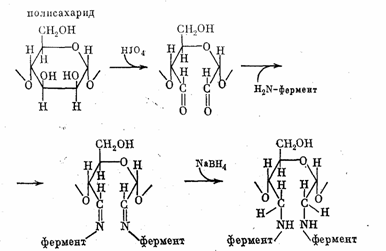
Рис. 1. Ковалентное присоединение фермента к полисахаридному носителю.
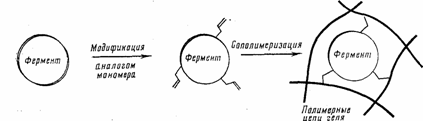
Рис. 2. Сополимеризационный метод для иммобилизации фермента в полимерном геле.
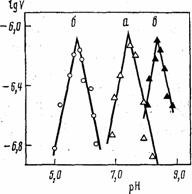
Рис. 3. рН-зависимость максимальной скорости гидролиза бензилпенициллина пенициллинамидазой.
а - нативный фермент; комплекс фермента с поликатионом (б) и полианионом (в).
Ферменты подвержены опасности «поедания» их микроорганизмами, почти всегда присутствующими в окружающей среде. Этого легко избежать, если фермент экранировать от них, включив его, например, в микропористый носитель. В результате связывания фермента с носителем становятся невозможными (или хотя бы затрудненными) также и различного рода полимолекуляряые инактивационные процессы типа фермент-ферментного взаимодействия, такие, как агрегация или автолиз в случае протеолитических ферментов.
Другой весьма распространенный механизм инактивации ферментов [6] состоит в их денатурации при неблагоприятных условиях внешней среды (повышенная температура, крайние значения рН, органические растворители). Денатурационному разворачиванию ферментной молекулы удается воспрепятствовать, если каталитически активную конформацию искусственно закрепить. Идея некоторых наиболее оправдавших себя принципов представлена на рис. 4 пространственное строение белковой молекулы становится более устойчивым при наложении на нее скобок (проводя внутримолекулярную «сшивку» белка бифункциональными реагентами, см. рис. 4, а), в результате многоточечного ковалентного или нековалентного присоединения фермента к носителю (рис. 4, б) или при механическом включении его в «тесные» поры носителя (рис. 4, в). Таким путем денатурацию (инактивацию) белка удалось замедлить в сотяи, тысячи и миллионы раз (в зависимости от метода стабилизации и природы фермента). Это означает, что, фермент, который, например, при температуре 50°С инактивируется практически мгновенно (рис, 5), будучи стабилизованным, может «работать» при температуре 80° часами или сутками и при более низкой температуре—месяцами.
Удалось также решить задачу как «заставить» ферменты функционировать в органических растворителях с малым содержанием воды. С этой целью применяют двухфазные системы вода - несмешивающийся с водой органический растворитель [16]. Гетерогенную реакционную среду создают обычно как микроэмульсию водного раствора фермента в органической среде или, что гораздо более удобно методически и технологически, как суспензию в органическом растворителе пористых частиц (стекло, керамика и др.), пропитанных водным раствором фермента (рис. 6, а). Весьма важная разновидность подобного рода реакционных сред - это коллоидный раствор воды в органическом растворителе [14]. Здесь фермент, будучи включенным в обращенную мицеллу поверхностно-активного вещества (детергента, липида), функционирует в своеобразном микрореакторе, содержащем всего лишь несколько сот или тысяч молекул воды (рис. 6,6).
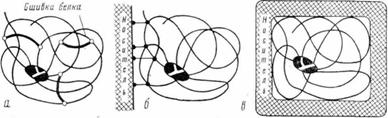
Рис. 4. Физико-химические принципы, позволяющие закрепить структуру ферментной, глобулы
а - внутримолекулярная «сшивка»; б - ковалентное или нековалентное присоединение к носителю; в - механическое включение в «тесные» поры носителя
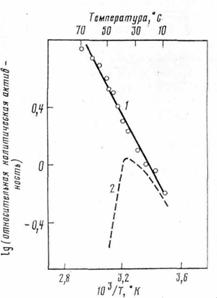
Рис. 5. Температурная зависимость относительной каталитической активности
1 - химотрипсин, иммобилизованный в по-лиакриламидном геле по сополимеризаци-опной методике (см. рис. 2); 2—свободный фермент [37]
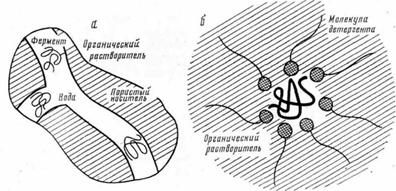
Рис. 6. Двухфазная система «вода - несмешивающийся с водой органический растворитель» с ферментом, включенным в поры носителя (а), и включение фермента в обращенные мицеллы поверхностно-активного вещества в органическом растворителе (б)
Наряду с созданием научных основ стабилизации ферментов сейчас успешно решаются также и другие проблемы, связанные с продлением их работоспособности, такие, как реактивация денатурированных ферментов и регенерация кофакторов [16].
Основные области применения иммобилизованных ферментов. Особенно ощутимый вклад иммобилизованные ферменты внесли в тонкий органический синтез, в анализ, в медицину, в процессы конверсии энергии, в пищевую и фармацевтическую промышленность.
Органический синтез [15]. Применение ферментов в органическом синтезе сдерживалось до последнего времени прежде всего тем, что многие органические реагенты плохо растворимы или вообще нерастворимы в воде как классической среде ферментативных реакций. Более того, положение равновесия многих практически важных процессов таково, что нужные продукты могут быть получены с высоким выходом лишь при проведении реакции в органическом растворителе. В этих условиях, однако, ферменты теряют каталитическую активность. Поэтому создание научных основ двухфазного метода (см. рис. 6) послужило решающим стимулом в развитии ферментативного синтеза. Для синтетической органической химии важно то, что в двухфазных реакционных средах фермент сохраняет каталитическую активность даже при исключительно малом содержании воды (доли процента) и, следовательно, равновесие катализируемой реакции (выход продукта) экспериментатор может регулировать в широких пределах, подбирая нужный, т. е. подходящий для синтеза, органический растворигель.
Иммобилизованные ферменты дали толчок к созданию принципиально новых методов «безреагентного» непрерывного анализа многокомпонентных систем органических (а в ряде случаев и неорганических) соединений. Главное звено в этих методах—«ферментные электроды» и «ферментные термисторы». В будущем важную роль в контроле окружающей среды и в ( клинической диагностике должны сыграть такие методы, как биолюминесцентный анализ [8] и иммуноферментный анализ.
Иммобилизованные ферменты в медицине открыли, во-первых, путь к созданию лекарственных препаратов пролонгированного действия со сниженной токсичностью и аллергентностью [9]. В СССР был создан первый в мире препарат иммобилизованного фермента (стрептокиназа) для парентерального введения при лечении сердечно-сосудистых заболеваний [8, 3]. Во-вторых, иммобилизационные подходы способствуют решению проблемы направленного транспорта лекарств в организме [2].
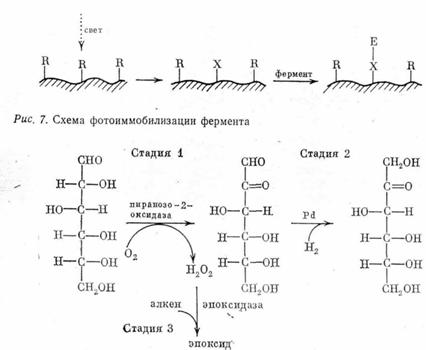
Рис. 8. Технологический процесс совместного получения фруктозы и алкен-оксидов.
Биоконверсия энергии и массы. Эти задачи пытаются успешно решать прежде всего микробиологическим путем. Тем не менее иммобилизованные ферменты вносят ощутимый вклад в осуществление биофотолиза воды и в биоэлектрокатализ [6, 8, 10], и с их участием становится реальным создание топливных элементов [10]. Заслуживает внимания также проблема деструкции возобновляемого растительного сырья, прежде всего расщепление лигниноцеллюлозных материалов [8].
Искусственные ферментативные усилители слабых сигналов. На активный центр иммобилизованного фермента удается действовать через носитель, подвергая последний механическим нагрузкам, ультразвуковой обработке или фотохимическим превращениям. Это позволяет регулировать каталитическую активность системы фермент—носитель под действием механических [5], ультразвуковых [9] и световых [7, 17] сигналов. На этой основе были созданы механо- и звукочувствительные датчики [9] и открыт путь к бессеребряной фотографии [13].
Идея как получить изображение с помощью метода иммобилизации ферментов, исключительно проста (рис. 7). функциональные группы исходного носителя (подложки) не должны взаимодействовать с ферментом, однако под действием света они претерпевают химическое превращение (К-»Х) с образованием фрагментов, у которых появилась способность химически связывать фермент или его сорбировать. Таким образом, фермент оказывается иммобилизованным на поверхности подложки именно в тех местах, куда попал свет, а в неосвещенном участке его не будет. Иными словами, в результате фотоиммобилизации фермента возникает «скрытое изображение». «Пришитый» фермент можно визуализовать, если подложку обработать раствором субстрата, который под действием света превращается в окрашенный продукт. В принципе при проявлении подложки субстратом каждая молекула фотоиммобилизованного фермента нарабатывает миллионы молекул окрашенного продукта (каталитический эффект усиления первичного сигнала), тем самым скрытое изображение становится видимым невооруженным глазом. В отличие от других ферментативных фотографических процессов [3] фотоиммобилизационный метод [27] весьма универсален, поскольку реализация его фактически не зависит от природы фермента.
Промышленные процессы. Технологические процессы с применением иммобилизованных ферментов внедрены прежде всего в пищевую и фармацевтическую промышленность. Главная мысль, которую нам хотелось бы здесь подчеркнуть, заключается в том, что биокатализ начинает стирать привычные границы между пищевыми и органосинтетическими технологиями. Наглядный пример — процесс совместного получения фруктозы и окислов алкенов, который разрабатывает фирма “CETUS Corporation” в США (рис. 8). На первой стадии проводят окисление D-глюкозы до D-глюкозона под действием иммобилизованной пиранозо-2-оксидазы; на второй стадии под действием водорода на палладиевом катализаторе полученный глюкозой восстанавливают со 100%-ным выходом до D-фруктозы. Побочный продукт первой стадии (перекись водорода) D-используется для микробиологического окисления этилена или пропилена до соответствующих эпоксидов. Как видно, в данном процессе слились три синтетических метода: ферментативный (стадия 1), химический (стадия 2) и микробиологический (стадия 3). Этот процесс, наполовину пищевой (фруктоза), наполовину органосинтетический (эпоксиды), можно считать предвестником будущих, еще более совершенных, т. е. комплексных и безотходных, биотехнологий.
Применение иммобилизованных ферментов в пищевой промышленности
Использование биологических процессов и агентов для получения пищевых продуктов и улучшения их качества - древнейшая ветвь биотехнологии. Скорее, это ее корни. Примеры в этом отношении общеизвестны — получение молока, изготовление вин, уксуса, пивоварение, сыроделие, хлебопечение и т. д. Во всех этих процессах в качестве биологических агентов выступали и продолжают выступать биологические организмы - от диких и домашних животных до микроорганизмов.
История пищевых технологий насчитывает тысячелетия, и тем не менее совершенствование их продолжается не только неослабевающими, но даже возрастающими темпами. Масштабы современных исследований и разработок в области традиционных технологий пищевых продуктов огромны. Однако в последнее время наметились перспективы принципиального сдвига в технологиях получения и улучшения качества, пищевых продуктов: разработки в этой области стали переходить от использования целых биологических организмов на клеточный и молекулярный уровни [5].
Как известно, все живые организмы содержат большое количество (сотни и тысячи) ферментов, основная функция которых состоит в проведении, ускорении и регуляции практически всех химических реакций, необходимых для жизнедеятельности организма. Благодаря высокой активности и специфичности некоторые ферменты уже давно нашли применение в ряде областей промышленности:
Ферменты Области использования
Гликозидазы
![]() -амилаза Хлебопечение
-амилаза Хлебопечение
» Пивоварение
» Производство кондитерских изделий
» Обработка текстильных изделий
» Производство высококачественной бумаги
глюкоамилаза Получение глюкозы
» Осахаривание ликеров и пива
инвертаза Производство кондитерских изделий лактаза Производство мороженого пектиназы Производство и осветление вин и фруктовых соков
» Производство кофе
целлюлазы Облагораживание растительных кормов
» Приготовление морковных джемов
» Обработка цитрусовых
Протеазы
протеазы микробного
происхождения Размягчение мяса
» Добавки к детергентам
» Обработка текстильных изделий
» Хлебопечение
» Осветление вин и пива
» Стабилизация сгущенного молока
» Выделка кож
бромелаин Размягчение мяса
» Стабилизация пива
папаин Осветление пива
» Получение белковых гидролизатов молока
» Размягчение мяса
фицин Размягчение мяса
трипсин Выделка кож
реннин Сыроделие
пепсин Сыроделие
» Стабилизация пива
Липазы Модификация вкуса молочных продуктов
» Сыроделие
Оксидоредуктазы Производство кондитерских изделий
» Выделка кож
глюкозооксидаза Удаление кислорода и/или глюкозы из пи-
щевых продуктов
» Пивоварение
» Приготовление майонезов
каталаза Удаление перекиси водорода после стерили-
зации молочных продуктов, после отбелива-
ния тканей, кожи,древесины и т. д.
Изомеразы
глюкозоизомераза Производство глюкозо-фруктозных сиропов
В основном, как видно, это относится к пищевой промышленности, где применяются главным образом малоочищенные комплексные ферментные препараты для гидролиза природных полимеров — белков, крахмала, пектинов.
В последние полтора-два десятилетия определились пути расширения областей применения ферментов. Они связаны с получением так называемых иммобилизованных ферментов, а также иммобилизованных клеток микроорганизмов [2]. Сочетание уникальных каталитических свойств ферментов с их водонерастворимостью в иммобилизованном виде послужило основой для создания ряда новых технологических процессов [4]. Эти процессы в настоящее время применяются главным образом в производстве пищевых продуктов.
К настоящему времени шесть процессов с использованием иммобилизованных ферментов или клеток нашли крупномасштабное применение в пищевой промышленности ряда развитых стран мира.
1. Получение глюкозо-фруктозных сиропов и далее фруктозы из глюкозы.
2. Получение оптически активных D-аминокислот из их рацемических смесей.
3. Синтез L-аспарагиновой кислоты из фумаровой кислоты.
4. Синтез L-яблочяой кислоты из фумаровой кислоты.
5. Получение диетического безлактозного молока.
6. Получение cахаров из молочной сыворотки.
Получение глюкозо-фруктозных спиртов
Фруктоза, или иначе фруктовый, плодовый или медовый сахар, широко распространена в природе. Этот особый сахар содержится во многих фруктах и плодах. Особенно богаты им яблоки и помидоры, а также пчелиный мед, который почти наполовину состоит из фруктозы. По сравнению с обычным сахаром (в состав молекул которого фруктоза также входит, но в виде химического соединения с менее сладкой глюкозой) фруктоза обладает более приятным вкусом, и, согласно профессиональной терминологии, вкус фруктозы «медовый», а обычного сахара—«приторный». Далее, по сладости фруктоза на 60-70% слаще сахара, и соответственно ее меньшее потребление влечет за собой меньшую калорийность продукта. Это важно с точки зрения диетологии питания. Наконец, фруктозу в отличие от глюкозы или сахара могут потреблять больные диабетом, и вообще замена сахара фруктозой существенно снижает вероятность этого заболевания. Дело в том, что физиологический путь использования человеческим организмом фруктозы совершенно другой, чем сахара или глюкозы, и не связан с превращением инсулина. К этому остается добавить, что фруктоза значительно менее вредна для зубов (из-за кариеса), чем сахар, и в смеси с глюкозой не кристаллизуется (не засахаривается), что важно в производстве мороженого, кондитерских изделий и т. д.
Несмотря на все эти неоспоримые преимущества фруктозы по сравнению с обычным сахаром, ее производство в мире практически отсутствовало вплоть до начала 70-х годов. Однако после того как в 1973 г. американской компанией «Клинтон Корн» был внедрен в промышленность процесс превращения глюкозы во фруктозу под действием иммобилизованного фермента глюкозоизомеразы, этот процесс стал самым крупным в мире по сравнению с другими, в которых используются иммобилизованные ферменты.
Научные основы процесса довольно просты. Фермент глюкозо-изомераза-катализирует превращение (изомеризацию) глюкозы во фруктозу в одну стадию, и реакция протекает до тех пор, пока в реакционной системе количества глюкозы и фруктозы не станут приблизительно равными. После этого реакция останавливается, и полученную смесь можно либо использовать в виде глкжозо-фруктозного сиропа, либо отделить фруктозу и оставшуюся глюкозу опять подвергнуть изомеризации. Этот процесс проводят непрерывно в реакционных колоннах высотой до 5 м, - которые предварительно заполняют иммобилизованным ферментом в виде гранул, полых нитей, кусочков геля (типа желатины) и т. д. В колонну непрерывным потоком подают раствор глюкозы (предварительно полученной при гидролизе кукурузного или картофельного крахмала), из колонны вытекает глюкобзо-фруктозный сироп.
Об эффективности такой технологии свидетельствуют следующие данные: на 1 кг иммобилизованного фермента за 100 дней работы получается 4 т фруктозы (в пересчете на сухой продукт) [4]. При этом время, за которое активность фермента уменьшается вдвое (так называемое время полуинактивации фермента) составляет от 20 до 50 дней. Иначе говоря, катализатор (иммобилизованный фермент) в колонне следует заменять только раз в 2 или даже 3 мес и процесс благодаря этому становится экономически выгодным, особенно по сравнению с использованием растворимой глюкозоизомеразы. Расчеты показывают, что в последнем случае затраты на фермент в 10 раз выше, и трудозатраты выше в 3 раза. В целом стоимость процесса с иммобиллизованным ферментом составляет лишь 61% от стоимости процесса с растворимым ферментом [4].
Современный завод по производству глюкозо-фруктозных сиропов с помощью иммобилизованного фермента, которых в мире имеется уже несколько десятков, производит до 400 т продукта в день в пересчете на сухое вещество: В целом мировое производство глюкозо-фруктозных сиропов в 1980 г. достигло 2,5 млн. т и, по оценкам, составит около 4 млн. т в 1985 г. и Свыше 5 млн. т в 1990 г. В США уже в 1998 г. потребление населением фруктозы составило 12% от потребления сахара и ожидается дальнейший рост вплоть до 30—40% к 2000 г. В Японии уже к 1985—1990 г,. около 30% потребляемого в стране сахара будет заменено на глюкозо-фруктозную смесь и чистую фруктозу [4].
9.2. Получение аминокислот
Растения и микроорганизмы способны сами синтезировать все нужные им аминокислоты из более простых химических соединений. Однако человеческий организм способен синтезировать лишь 12 из 20 так называемых «магических» аминокислот, необходимых ему для жизнедеятельности. Остальные 8 аминокислот получили название незаменимых и должны поступать в организм извне - с пищей. При нехватке хотя бы одной из незаменимых аминокислот замедляется рост организма, начинаются болезни. Поэтому важно производить эти аминокислоты для корректировки рационов питания, в лечебных и профилактических целях и т. д.
Производство многих аминокислот, в том числе и незаменимых крупнотоннажная отрасль химической промышленности. Однако химики производят смесь оптических изомеров аминокислот, иначе говоря, смесь L- и D-аминокислот, молекулы которых в L- и D-форме представляют собой зеркальные отражения друг друга. В химических реакциях эти изомеры практически неразличимы, более того, по многим химическим критериям смесь L- и D-изомеров аминокислот представляет собой «химически чистое» вещество. Однако в большинстве случаев (исключением является лишь метионин) человеческий организм усваивает лишь L-аминокислоты, а их D-изомеры являются нежелательной примесью.
Разделение смеси L- и D-аминокислот, так называемой рацемической смеси, на составляющие ее изомеры стало первым процессом в мире, осуществленным с помощью иммобилизованных ферментов на промышленном уровне. Этот процесс был реализован в Японии компанией «Танабе Сейяку» в 1969 г. В течение 15 предшествующих лет данный процесс проводился компанией с применением растворимого фермента—аминоацилазы, но он был недостаточно экономичен. После перевода на иммобилизованную аминоацилазу экономическая эффективность процесса возросла в полтора раза, и в настоящее время компания осуществляет на промышленном уровне производство пяти L-аминокислот, из них четыре незаменимые (метионин, валин, фенилаланин, триптофан).
В качестве исходного вещества используются ацилированные L- и D-аминокислоты, полученные с помощью обычного химического синтеза, и их подвергают воздействию аминоацилазы. Фермент гидролизует только ацил-L-изомер, отщепляя от него объемную аыильную группу и тем самым резко увеличивая растворимость образующейся L-аминокислоты по сравнению с присутствующим в реакционной системе ацил-D-изомером. После этого вещества легко отделяются друг от друга с помощью известных физико-химических приемов, продуктом является чистая L-аминокислота.
Остающаяся ацил-D-аминокислота при нагревании рацемизуется, т. е. переходит опять в смесь ацилированных L- и D-аминокислот, и процесс повторяют сначала. Таким образом, в итоге единственным продуктом является L-аминокислота. Оказалось, что для аминоацилазы не имеет значения, какую аминокислоту ей гидролизовать, важно лишь строение ацильной части, к которой фермент имеет строгую специфичность. Благодаря этому одна и та же реакционная колонна с иммобилизованной аминоацилазой может быть использована для производства самых различных, L-аминокислот.
Иммобилизованный фермент легко готовить, так как он просто адсорбируется на специальной смоле, которую затем помещают в колонну объемом 1 м3. Время полуинактдвации иммобилизованного фермента в промышленных условиях составляет 65 дней, и, когда активность катализатора падает ниже нормы (раз в несколько месяцев), в колонну добавляют раствор свежего фермента, который опять адсорбируется на носителе. Устойчивость полимерного носителя для иммобилизации фермента столь высока, что он используется японской компанией более 8 лет в той же колонне без замены.
9.3. Получение L-аспарагиновой кислоты
Аспарагиновая кислота не принадлежит к числу незаменимых, но производится в мире многими тысячами тонн. Она находит широкое применение в пищевой промышленности для придания (в сочетании с другой аминокислотой—глицином) кондитерским изделиям и напиткам различных оттенков кислого или сладкого вкуса.
Замечателен процесс получения аспарагиновой кислоты с помощью фермента аcпартазы. С практической стороны он отличается тем, что в качестве исходных веществ для ферментативного синтеза используются фумаровая кислота и аммиак, крупнотоннажные продукты органического и неорганического синтеза. В отношении самой реакции процесс интересен тем, что фермент здесь всего лишь в одну стадию присоединяет молекулу аммиака к двойной связи: фумаровой кислоты, сразу приводя к оптически активной L-аспарагиновой кислоте. Наконец, особенностью процесса является и, что в нем впервые в технологической практике были использованы иммобилизованные клетки микроорганизма, содержащие, фермент в его естественной микробиой оболочке. Этот процесс былразработаняпонской фирмой «Танабе Сейяку» в 1993 г.
Сначала японсйие технологи попытались использовать в качестве катализатора, микробную массу, состоящую из живых клеток микроорганизмов, содержащих внутриклеточный фермент, аопартазу. Однако время полуинактивации фермента в этих условиях у составляло всего 10 дней и было неприемлемым для технологии. Когда аспартазу выделяли из клеток и иммобилизовали в геле, время полуинактивации увеличилось до 30 дней. Когда же в гель включили сами микробные клетки и дополнительно усилили их путем химического связывания друг с другом и с гелем, стабильность катализатора резко возросла и время полуинактивации фермента составило уже 120 дней. Плотный гель с иммобилизованными в нем микробными клетками, содержащими аспартазу, формуют в кубики размерами 2—3 мм, набивают ими колонну объемом 1 м3 и пропускают через нее раствор фумарата аммония. На выходе из колонны L-аспарагиновую кислоту кристаллизуют, центрифугируют и промывают холодной водой. Процесс практически полностью автоматизирован и осуществляется в непрерывном режиме. Масштабы производства на фирме «Танабе Сейяку»—1700, кг чистой L-аспарагиновой кислоты в день на реактор объемом 1 м3.
9.4. Получение L-яблочной кислоты
Яблочная кислота находит спрос на мировом рынке в качестве заменителя лимонной кислоты в продуктах питания и фармацевтических препаратах. Химическим путем (гидролизом ангидрида яблочной кислоты) производят только рацемическую смесь оптических изомеров яблочной кислоты, в то время как оптически активный L-изомер, который получают микробиологическим способом, пока слишком дорог для широкого промышленного производства.
В то же время L-яблочную кислоту можно получать ферментативным путем из той же фумаровой кислоты, из которой получают L-аспарагиновую кислоту (см. выше). Только в данном случае ферментом является фумараза, которая присоединяет по двойной связи фумаровой кислоты не аммиак, а воду. В остальном реакция происходит так же, и в качестве ферментного катализатора используют клетки, содержащие фумаразу и иммобилизованные в гель. В обычных (интактных) клетках время полуинактивации фумаразы составляет 6 дней, в иммобилизованных в полиакриламид-ный гель — 55 дней.
Недавно компания «Танабе Сейяку» изменила технологию и перешла к другому микробному источнику фумаразы, стабильность и продуктивность которого выше в два раза по сравнению с предыдущим. Помимо этого, компания стала использовать вместо поли-акриламидного геля в качестве носителя для иммобилизованных клеток другой гель на основе каррагинана — полисахарида из морских водорослей. В итоге время полуинактивации иммобилизованного фермента составило 160 дней и продуктивность его действия возросла в 5,2 раза по сравнению с прежней технологией.
9.5. Получение без лактозного молока
Лактоза, или молочный сахар, содержится в достаточно больших количествах в молоке и молочной сыворотке. Этот сахар характеризуется малой сладостью и низкой, растворимостью, и именно из-за его присутствия происходит кристаллизация мороженого и других молочных изделий и продуктов, которая иногда случается и приводит к малоприятным вкусовым ощущениям.
Молекулы лактозы состоят из глюкозы и галактозы и распадаются на них при гидролизе под действием лактазы или b-галактозидазы. Молоко после такой обработки приобретает новые диетические качества, поскольку определенная часть населения не может употреблять молоко именно из-за наличия в нем лактозы. Это качество организма получило название лактазной недостаточности, и в целом по направлению с севера нашей планеты на юг доля людей, испытывающих лактазную недостаточность заметно возрастает. В Африке, например, целые племена и этнические группы не могут пить молоко домашних животных из-за сильных аллергических откликов или неприятных физиологических ощущений. В то же время они нормально усваивают молоко, не содержащее лактозы или предварительно обработанное иммобилизованной лактазой.
Первый коммерческий процесс получения безлактозного молока с использованием иммобилизованной лактазы был осуществлен итальянской фирмой «Сентрале дель Латте» в Милане. Получаемое диетическое молоко несколько слаще обычного, поскольку глюкоза более сладкая, чем лактоза, однако это не мешает его употреблению. Стабильность иммобилизованного фермента достаточно высока, и после 50 дней работы он сохраняет 80% первоначальной активности. В настоящее, время завод в Милане производит 10 т безлактозного молока в день.
9.6. Получение сахаров из молочной сыворотки
Молочная сыворотка содержит в своем составе большое количество лактозы—около 5% в жидкой и 75% в высушенной сыворотке. Ферментативный гидролиз лактозы в этом случае открывает новые возможности получения сахаристых веществ и.з нетрадиционного сырья, вносит определенный вклад в решение кормовой проблемы и в проблему охраны окружающей среды, поскольку сыворотка большей частью не утилизируется.
Первый промышленный процесс гядролиза лактозы в молочной сыворотке с помощью иммобилизованной лактазы был реализован в 1980 г. совместно английской, французской и американской организациями одновременно в Англии и Франции.
Перед введением в колонный реактор с иммобилизованным ферментом сыворотку пастеризуют, подвергают ультрафильтрации и пропускают через ионообменник, таким образом ее деминерализуя. Мощность установки составляет около 1000 л/ч при степени конверсии лактозы 80%. Установка полностью автоматизирована. Получаемые при этом сахара (глюкоза и галаястоза) по сладости в полтора раза превышают сладость сахара в расчете на одинаковые экономические затраты.
Литература
1. Иммобилизованные ферменты Под ред. И. В. Березина, В. К. Антонова, К. Мартинека. М.: Изд-во МГУ, 1996. Т. 1. 296 с.; Т. 2. 358 с.
2. Березин И. В., Варфоломеев С. Д. Биокинетика. М.: Наука, 1979. 312 с.
3. Березин И. В., Варфоломеев С. Д., Казанская Н. Ф., Никольская И. И. Светочувствительные каталитические системы.—Усп. науч. фотографии, 1988, 19, с. 255-262.
4. Березин И. В. и др. Кинетические особенности катализа иммобилизованными ферментами.—Усп. химии, 1985.
5. Березин И. В. и др. Механохимия иммобилизованных ферментов.— Журн. физ. химии, 1975, 49, с. 2519—'2528.
6. Введение в прикладную энзимологию — иммобилизованные - ферменты/Под ред. И. В. Березина, К. Мартинека. М.: Изд-во МГУ, 1982. 383 с.
7. Березин И. В., Мартинек К. Искусственные ферментативные светочувствительные системы.—Усп. химии, 1989.
8. Химическая энзимология/Под ред. И. В. Березина, К. Мартинека. М.: Изд-во МГУ, 1983. 285 с.
9. Березин И. В., Мартинек К; Чазов Е. И. Ферментативные детекторы слабых сигналов.—Природа, 1980, № 11.
10. Варфоломеев С. Д. Конверсия энергии биокаталитическими системами. М.: Изд-во МГУ, 1981.256с.
11. Кулис Ю. Ю. Аналитические системы на основе иммобилизованных ферментов. Вильнюс: Мокслас, 1981. 200 с.
12. Марголин А. Л., Изумрудов В. А., Швядас В. Ю. и др. Обратимо растворимая пенициллинамидаза, иммобилизованная в полиэлектролитных комплексах—Докл. АН СССР, 1990.
13. Мартинек К„ Березин И. В. Стабилизация ферментов—ключевой фактор при внедрении биокатализа в практику.—Усп. химии, 1980, 49, с. 737—770.
14. Мартинек К; Левашов А. В., Клячко Н. Л., Березин И. В. Катализ водорастворимыми ферментами в органических растворителях.—Докл. АН СССР, 1977, 236, с. 920—923.
15. Мартинек К; Семенов А. Н. Катализ ферментами в органическом синтезе— Усп. химии, 1981, 50, с. 1376—1406.
16. Можаев В. В. Иммобилизация ферментов как подход к исследованию фундаментальных проблем биохимии.—Усп. биологической химии, 1983, 24.
17. Стрельцова 3. А., Швядас В. К; Максименко А. В. Влияние полиэлектролитов на свойства пенициллинамидазы,— Биоорган, химия, 1975.
