Використання методів ДНК-аналізу для діагностики моногенних спадкових захворювань
3. Результаты и обсуждения.
3.1. ДНК-анализ мутаций R408W, IVS12nt1, R158Q, Y414C, IVS10nt546 гена ФАГ.
На первом этапе мы изучали мутации R408W, Y414C (12-й экзон), R158Q (5-й экзон), Ivs10nt546 (10-й интрон), Ivs12nt1 (12-й интрон) гена ФАГ в семьях больных ФКУ из разных регионов Украины. Молекулярно-генетический анализ мутаций проводился согласно методике описанной в разделе 2.2.
Мутация R408W, локализована в 12-м экзоне и представляет собой единичную нуклеотидную замену цитозина на тимин, в результате чего триплет CGG, кодирующий аминокислоту аргинин в 408 положении заменяется на триплет TGG, кодирующий аминокислоту триптофан [74].
Общий принцип идентификации мупации R408W представлен на рисунке 3.1. Как видно из рисунка 3.1 и 3.2 при анализе мутация R408W мы наблюдали три набора рестрикционных фрагментов: 1) два фрагмента размером 148 пн. и 97 пн. свидетельствовали о наличии сайта рестрикции на обоих хромосомах, то есть о присутствии мутации в гомозиготном состоянии; 2) три фрагмента размером 245 пн., 148 пн. и 97 пн., соответствует сайту рестрикции на одной хромосоме - такая рестрикционная картина характерна гетерозиготным носителям мутации R408W; 3) один фрагмент размером 245 пн. соответствует сайтам рестрикции на обеих хромосомах что адекватно отсутствию, мутации R408W.

Рисунок № 3.1. Схема рестрикционного анализа для детекции мутаций R408W (эндонуклеаза рестрикции Eco1301).
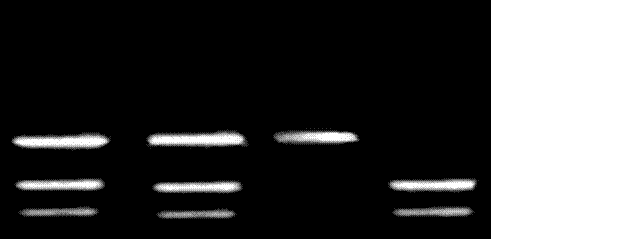
Рисунок. 3.2. ДНК - анализ мутации R408W гена ФАГ. 1.8 %-ной агарозний гель. 1- мать; 2- отец; 4- пробанд; 3- проба без мутации.
В результате проведенного исследования в 11 пробандов с 12 обследованных была выявленная мутация R408W. В шести пробандов мутация выявлена в гомозиготном состоянии, в пяти в гетерозиготном состоянии, в одного пробанда мутация R408W выявлена не была. ДНК-анализ родителей показал наличие мутации в гетерозиготном состоянии в 17 с 24 обследованных (Рис. 3.2.).
Таким образом, мутация R408W была выявлена на 34 с 48 хромосом 12 с мутациями гена ФАГ, то есть частота мутации R408W в проанализированной группе составляет 70 % от всех других мутаций. Полученные результаты свидетельствуют о том, что мутация была выявлена в 11 проанализированных семьях, то есть в 91.6 % от общего количества семей, причем в 50 % мутации были идентифицированы на обоих гомологичных хромосомах 12. Приблизительно в 41 % случаев мутация была идентифицирована на одной хромосоме из группы хромосом 12 и лишь в 9 % случаев мутации, которые служило причиной развития заболевания, остались не идентифицированными.
Для анализа мутации R158Q (CGG→САG, аргинин→глутамин), фрагмент 5-го экзона гена ФАГ амплифицировали in vitro с использованием праймера, вводящего единичную нуклеотидную замену в состав амплифицируемого фрагмента, в результате чего создается сайт рестрикции для эндонуклеазы MspI. При мутации R158Q сайт рестрикции исчезает (рис. 3.3 и 3.4) [70].
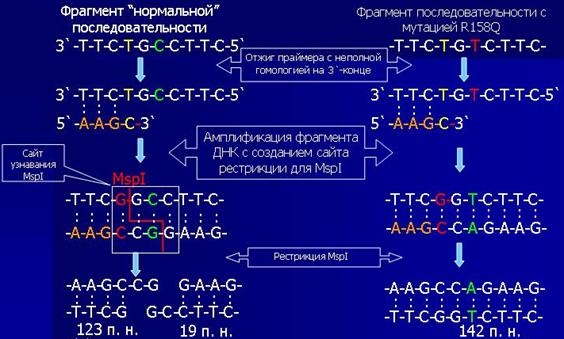
Рис. 3.3. Схема рестрикционного анализа для детекции мутаций R158Q (эндонуклеаза рестрикции MspI).
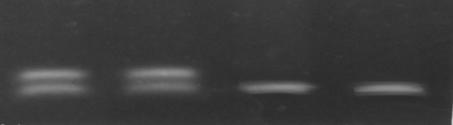
Рис. 3.4. Анализ мутации R158Q. 1,8 % агарозный гель. 1, 2 - R158Q/норма; 3 - норма/норма, 4 – котроль нома/норма.
Мутация R158Q была идентифицирована нами на 7-ми ФКУ хромосом. В шести случаях мутация находилась в компаунде с R408W и в одном случае в компаунде с неидентифицированной мутацией. Частота мутации R158Q составила 3,7%.
Мутация Y414C (TAC®TGC), представляет собой единичную нуклеотидную замену в 12-м экзоне гена ФАГ в результате, которой тирозин в 414-м положении заменяется на цистеин. Как мы видим при этой мутации происходит замена второго нуклеотида в составе аминокислотукодирующего триплета. Методика детекции этой мутации основана на использовании метода сайт-направленного мутагенеза, при котором в процессе ПЦР в состав амплифицируемого фрагмента посредством праймера с неполной гомологией на 3`-конце вводится единичная нуклеотидная замена, создающая сайт рестрикции для эндонуклеазы RsaI. При мутации этот сайт исчезает, что и является маркером мутации (рис. 3.5) [70].
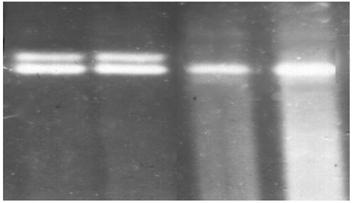
Рис. 3.5. Анализ мутации Y414C. 1,8% агарозный гель. 1, 2 – Y414C/норма; 3 – норма/норма, 4 – контроль, норма/норма.
В проанализированной нами группе мутация Y414C была выявлена у трех пробандов и во всех случаях в компаунде с мутацией R408W, а ее частота составила 1,6%.
Мутация IVS10nt546, замена G на А в положении 546 в 10-м интроне гена ФАГ, на расстоянии 11 п.н. от границы 10 интрон/11 экзон. В отличиe от всех предыдущих мутаций IVS10nt546 локализована в последовательности интрона и не влечет за собой аминокислотных замен, но приводит к нарушению сплайсинга. Нуклеотидная замена, возникающая при этой мутации, приводит к возникновению сайта для эндонуклеазы DdeI в последовательности 10-го интрона, что и служит маркером используемым при анализе мутации (рис. 3.6) [73].
Рис. 3.6. Анализ мутации Ivs10nt546. 1,8 % агарозный гель. 1, 2 – Ivs10nt546/норма; 3, 4 - норма/норма
В проанализированной нами группе три пациента были носителями данной мутации в компаунде с мутацией R408W. Частота мутации Ivs10nt546 в Украине составила 1,5 %.
Мутация IVS12nt1, представляет собой единичную нуклеотидную замену G на А в последовательности сайта сплайсинга в 12-ом интроне [75]. Также как и предыдущая мутация Ivs10nt546 Ivs12nt1 относится к группе мутаций сайтов сплайсинга. Маркером этой мутации является сайт рестрикции эндонуклеазы RsaI, который создается в «нормальной» амплифицируемой последовательности при участии праймера с единичной нуклеотидной заменой на 3`-конце и исчезает при мутации IVS12nt1 (рис. 3.7) [71].

Рис. 3.7. Анализ мутации Ivs12nt1. 1,8% агарозный гель. 1 - маркер молекулярной массы Ladder 100 bp , 2 – Ivs12nt1/норма; 3, 4 – норма/норма.
Нами было показано, что в популяции Украины мутация Ivs12nt1 встречается с частотой 1 %. Оба пациента, у которых была выявлена эта мутация, имели генотип R408W/Ivs12nt1.
3.1.2. ДНК-анализ мутаций в последовательности 7-го экзона гена ФАГ
На сегодняшний день в 7-м экзоне гена ФАГ идентифицировано 69 мутаций [37], поэтому следующим этапом нашей работы был скрининг мутантных вариантов именно в этом экзоне.
На основе анализа литературных данных нами были подобраны шесть наиболее распространенных мутаций последовательности седьмого экзона гена ФАГ: P281L, G272X, S273F, R261Q, R261X, R252W, которые было целесообразно протестировать у пациентов [82, 83]. Условия проведения полимеразной цепной реакции представлены в разделе 2.2.
В процессе проведения амплификации мы столкнулись с проблемой высокого выхода неспецифики, по этому мы были вынуждены пойти по пути оптимизации условий проведеия амплификации. В первую очередь была предпринята попытка увеличить температуру амплификации, с 55ºС до 57 ºС, а затем до 58ºС. При 57ºС неспецифика по прежнему присудствовала, а при 58ºС ПЦР продукт отсудствовал. Затем мы использовали многоступенчатый отжиг праймеров с элементами touch-down ПЦР:
1) предварительная денатурация 5 мин при температуре 94ºС;
2) 5 циклов каждый состоит из трех этапов (2.1 – 2.3);
2.1) денатурация ДНК – 94 ºС на протяжении 40 с;
2.2) отжиг праймеров - 59 ºС на протяжении 40 с.
2.3) отжиг праймеров - 61 ºС на протяжении 40 с.
2.4) элонгация 72 ºС на протяжении 50 с.,
3) 30 циклов каждый состоит из трех этапов (3.1 – 3.3);
3.1) денатурация ДНК – 94 ºС на протяжении 40 с;
3.2) отжиг праймеров - 57 ºС на протяжении 45 с.
3.3) отжиг праймеров - 59 ºС на протяжении 20 с.
3.4) отжиг праймеров - 60 ºС на протяжении 20 с.
3.5) элонгация 72 ºС на протяжении 50 с.,
4) элонгация 72ºС на протяжении 7 мин.
Амплификация с этим температурным циклом прошла, была получена высокая концентрация специфического ПЦР продукта при отсутствующей неспецифике.
Мутация R252W (CGG®TGG), приводит к синтезу белка, у которого аргинин, расположенный в 252-м положении, заменен на триптофан. Для детекции этой мутации ПЦР продукт 7-го экзона гена ФАГ обрабатывали эндонуклеазой Eco881, сайт рестрикции для которой исчезает при мутации (рис. 3.8) [76].

Рис. 3.8. Анализ мутации R252W. 1,8% агарозный гель. 1 – R252W/норма; , 2 – норма/норма. 3 - маркер молекулярной массы Ladder 100 bp.
Данная мутация была выявлена нами у 3-ти пациентов: в четырех случаях в компаунде с мутацией R408W и в одном случае в компаунде с неидентифицированной мутацией. Ее частота составила 2,9 %.
Аргинин в 261-м положении, образуя водородные связи с другими аминокислотами, выполняет функцию стабилизации структуры активного центра. Для этого аминокислотного остатка в последовательности белка известны три мутации R261Q (CGA→CAA, аргинин→глутамин), R261X (CGA→TGA, аргинин→стоп кодон) и R261P (CGA→CСA, аргинин→пролин). Детекцию первых двух представлялось возможным провести с использованием рестрикционного анализа в ходе данной работы. Маркером мутации R261Q служит сайт рестрикции для эндонуклеазы HinfI, который присутствует в «нормальной» последовательности 7-го экзона гена ФАГ и отсутствует при мутации R261Q [74]. Аналогичная ситуация с рестриктазой DdeI наблюдается при мутации R261Х [76].
Мутация R261Q была выявлена у двух пациентов. У одного пациента в компауде с мутацией R408W, а у второго в компаунде с неидетифицированной нами мутацией. Таким образом, нами было показано, что данная мутация встречается в Украине с частотой 1,2 %. Мутация же R261X выявлена не была.
Мутация S273F (TCC®TTC) приводит к нарушению структуры активного центра. При замене цистеина на тимин в положении 273 в последовательности 7-го экзона гена ФАГ формируется сайт рестрикции для эндонуклеазы Sau3A, что и служит маркером мутации, используемым для ее детекции [75]. Данная мутация была выявлена у одного пациента в компаунде с мутацией R413P. Распространена мутация S273F в Украине с частотой 0,6%.
Аминокислотный остаток пролина в 281-м положении является последней аминокислотой последовательности 7-го экзона. Кодируется данная аминокислота триплетом CCg, который при мутации P281L заменяется на триплет CTg, соответствующий лейцину. Для идентификации данной мутации ПЦР продукт 7-го экзона гена ФАГ, синтезированный с использованием метода сайт направленного мутагенеза, гидролизируют эндонуклеазой MspI, сайт узнавания для которой формируется в процессе ПЦР на «нормальной» последовательности гена ФАГ, но отсутствует при мутации P281L (рис 3.9) [77].

Рис. 3.9. Анализ мутации P281L. 1,8 % агарозный гель. 1 - P281L/норма; 2, 4 - норма/норма, 3 - маркер молекулярной массы Ladder 100 bp.
В проанализированной нами группе данная мутация была выявлена у двух пациентов. Во всех случаях в компаунде с мутацией R408W. Частота мутации P281L в исследуемой популяции составила 2,3 %.
Мутация G272X – относится к классу нонсенс-мутаций. Расположенный в 272-м положении гуанин заменяется на стоп кодон, что терминирует транскрипцию гена ФАГ и приводит к синтезу транскрипта у которого отсутствуют последние 6 экзонов. Для детекции мутации G272X ПЦР продукт 7-го экзона гена ФАГ гидролизируют эндонуклеазой BamHI, сайт рестрикции для которой отсутствует в мутантной последовательности.
3.1.3. Анализ алельного полиморфизма VNTR-локуса 3`-нетранслированной области гена ФАГ.
Анализ аллельного полиморфизма VNTR-локуса 3'- нетранслируемой области гена ФАГ проводили среди 470 здоровых доноров из разных регионов Украины (Киева, Львова, Кременчуга Полтавской области, Алчевска Луганской области). В результате исследования было выявлено 7 аллельных вариантов минисателитного VNTR-локуса. Число тандемных повторов при этом варьировало от 3 до 12, а размеры продуктов амплификации варьировали от 380 до 650 п.н. соответственно числу тандемных повторов. На рисунке 3.10 представленны некоторые аллельные варианты VNTR-локуса.
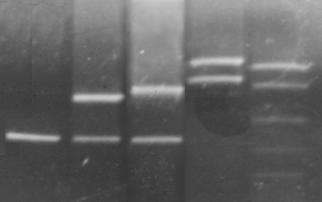
Рис. 3.10. Электрофореграмма продуктов амплификации минисателлитного VNTR –полиморфизма, 1.8% агарозний гель: 1 - генотип 3/3 (380/380 п.н.);2 - генотип 3/7 (380/500 п.н.); 3 - генотип 3/8 (380/530 п.н.); 4 - генотип 9/12 (560/650 п.н.); M - Маркер молекулярной массы ДНК плазмиды pBR322 гидролизированная эндонуклеазой MspI.
3.2. Оптимизпция условий проведения амплификации.
Объектом оптимизации может выступоть тест-система для диагностики фенилкетонурии, а также другие тест системы.
Основной проблемой при амплификации ДНК in vitro является не специфика. Специфичность амплификации – это отношение концентрации целевого ПЦР продукта ко всему ПЦР продукту.
На специфичность амплификации определенной последовательности ДНК влияют следующие факторы:
1) Специфичность отжига праймеров.
2) Ошибка в работе полимеразы.
3) Полнота денатурации цепей участка ДНК матрицы во время удлинения цепи.
Основным направлением оптимизации амплификации является увеличение специфичности отжига праймеров при высокиоких значениях выхода продукта.
Специфичность отжига праймеров.
Отжиг праймеров может проходить в различных локусах ДНК матрицы. При выборе праймеров выбирают такие праймеры, которые имеют только один уникальный участок отжига на матрице. Как будет показано ниже, праймеры могут отжигаться и на не полностью комплементарной матрице.
Отжиг праймеров на не полностью гомологичной матрице представлен на рисунке 3.1, где первые семь проб - ПЦР с одной парой праймеров, вторые семь проб - мультиплексная ПЦР. Разная интенсивность ПЦР продукта может быть объяснена тем, что при амплификации с одной парой праймеров нет конкуренции праймеров за ssДНК, а при мультиплексной ПЦР наблюдается конкуренция между праймерами за ssДНК. Эта конкуренция приводит к тому, что при мультиплексной ПЦР приймущественно удлиняются комплексы праймер-ssДНК, которые обладают максимальной гомологией и, как следствие, - максимальной устойчивостью и концентрацией.
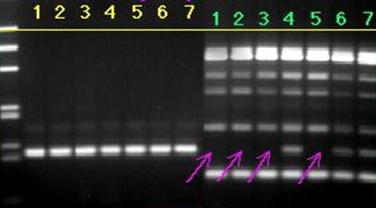
Рисунок № 3.1. Продукты ПЦР, полученные при одинаковых условиях ПЦР – цикла на семи разных матрицах. Первые семь проб ПЦР - с одной парой праймеров, вторые семь проб - мультиплексная ПЦР.
Это может быть особенно актуально, когда отжигу праймеров с целевым участком препятствует стерический фактор или другие факторы. В таких случаях увеличивают температуру и время отжига праймеров. Однако увеличение температуры отжига праймеров приводит к снижению интенсивности амплификации (см. рисунок № 1.5.), что обусловлено снижением разности энергии Гибса в реакции образования ДНК.Pr и, как следствие, - устойчивости комплекса ДНК.Pr.
В связи с этим исследователи проводят много ступенчатый отжиг праймеров при разной температуре. При этом они полагаются как на собственный опыт, так и на опыт коллег. И так, если не удается провести отжиг праймеров при одной температуре, проводят многоступенчатый отжиг праймеров. При многоступенчатом отжиге праймеров, как правило, температуру увеличивают см. таблицу № 1, иногда проводят многоступенчатое снижение температуры.
Таблица № 1.
| Температура |
Время |
| 56 С0 |
40 c |
| 57 С0 |
20 c |
| 58 С0 |
20 c |
| 60 С0 |
30 c |
Целью раздела 3.1. будет рассмотрение преимуществ и недостатков многоступенчатого отжига праймеров.
3.1. Постановка задачи оптимизации.
При постановке амплификации у нас есть два критерия оценки ее эффективности: выход продукта и специфичность. Мы заинтересованы в максимизации обоих критериев. Основным инструментом для изменения этих критериев является температура отжига праймеров. Однако любое изменение температуры приводит к увеличению одного из параметров и снижению другого. Возможны следующие варианты изменения температуры во времени, которые мы рассмотрим далее:
- ступенчато изменяющаяся температура;
- температура, заданная в виде функции:
- равномерно уменьшающаяся (увеличивающаяся) температура, функция (3.85);
- функции (3.86-3.88).
Для выбора оптимального режима изменения температуры отжига праймеров необходимо задать начальные ограничения: специфичность одного раунда амплификации выше 99%, процент отжогшихся праймеров выше 75%.
3.2. Вывод математической модели амплификации.
Для оценки преимуществ и недостатков многоступенчатого отжига праймеров необходимо вывести математическую модель амплификации.
Для этого рассмотрим конкурентные процессы, которые происходят при отжиге праймеров.
Выделим четыре процесса, протекающие при отжиге праймеров, и проанализируем их влияние на специфичность амплификации.
1) Образование комплекса ДНК.Pr
ДНК+Pr![]() ДНК.Pr (3.1)
ДНК.Pr (3.1)
Скорость данного процесса, согласно закона действующих масс, описывается уравнением (3.2).
V1=![]() (3.2)
(3.2)
где k1 – константа скорости образования комплекса ДНК.Pr. Так как процесс проходит со значительным избытком Pr, то C(Pr) можно вести в константу, тогда уравнение примет вид (3.3).
V1=![]() (3.3).
(3.3).
2) Разрушение комплекса ДНК.Pr:
ДНК.Pr![]() ДНК+Pr (3.4)
ДНК+Pr (3.4)
Скорость данного процесса, согласно закона действующих масс, описывается уравнением (3.5).
V2=![]() , (3.5)
, (3.5)
где k2 – константа скорости разрушения комплекса ДНК.Pr.
3) Образование комплекса ДНК*.Pr с неполной гомологией.
ДНК+Pr![]() ДНК*.Pr (3.6)
ДНК*.Pr (3.6)
Скорость данного процесса, согласно закона действующих масс, описывается уравнением (3.7).
V3=![]() , (3.7)
, (3.7)
где k3 – константа скорости образования комплекса ДНК*.Pr. Так как процесс проходит с значительным избытком Pr, то C(Pr) можно вести в константу, тогда уравнение примет вид (3.8)
V3=![]() (3.8).
(3.8).
4) Разрушение комплекса ДНК*.Pr с неполной гомологией.
ДНК*.Pr![]() ДНК+Pr (3.9)
ДНК+Pr (3.9)
Скорость данного процесса, согласно закона действующих масс, описывается уравнением (3.10).
V4=![]() , (3.10)
, (3.10)
где k4 – константа скорости разрушения комплекса ДНК*.Pr.
Применив закон сохранения массы, получим уравнение (3.11), продифференцировав которое получим уравнение (3.12).
С(ДНК)+![]() +
+![]() = С(ДНК)0 (3.11)
= С(ДНК)0 (3.11)
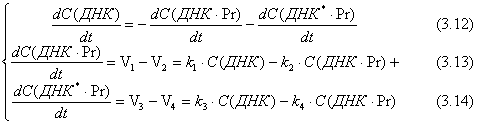
Для простоты записи заменим x = С(ДНК), y = C(ДНК.Pr), z = C(ДНК*.Pr), a = C(ДНК)0 и получим:
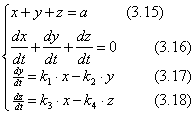
В данном уравнении k1, k2, k3, k4 = const.
Подставим уравнение (3.18) в уравнение (3.16) и получим:
![]() (3.19)
(3.19)
Выразим z из уравнения (3.15) и подставим в (3.19):
![]() (3.20)
(3.20)
![]() (3.21)
(3.21)
Выразим x и ![]() из уравнения (3.17) и получим уравнения (3.22) и (3.23).
из уравнения (3.17) и получим уравнения (3.22) и (3.23).
![]() (3.22)
(3.22)
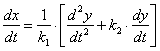 (3.23)
(3.23)
Подставивши x из уравнения (3.22) и ![]() из уравнения (3.23) в уравнение (3.21) получим уравнение (3.24)
из уравнения (3.23) в уравнение (3.21) получим уравнение (3.24)
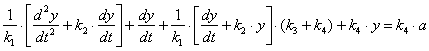 (3.24)
(3.24)
![]()
![]() (3.25)
(3.25)
![]()
![]() (3.26)
(3.26)
Уравнение (3.26) это линейное не однородное дифференциальное уравнение второго порядка. Его общее решение можно представить как сумму общего решения однородного уравнения yоо и частичного решения неоднородного уравнения yчн.
Однородное уравнение имеет вид:
![]() (3.27)
(3.27)
Решение однородного уравнения второго порядка необходимо искать в виде ![]() .
.
Найдем первую и вторую производную общего решения однородного уравнения:
![]() (3.28)
(3.28)
![]() (3.29)
(3.29)
Подставивши выражения (3.28), (3.29) в выражение (3.27), получим выражение (3.30).
![]() (3.30)
(3.30)
Учитывая, что ![]() , получим выражение (3.31).
, получим выражение (3.31).
![]() (3.31)
(3.31)
Уравнение (3.31) является квадратным уравнением, решением которого будет уравнение (3.32).
![]() (3.32)
(3.32)
Найдя корни уравнения (3.31), мы нашли корни дифференциального уравнения второго порядка с однородной правой частью.
![]() (3.33)
(3.33)
Так как левая часть уравнения (3.26) константа, то частичное решение неоднородного уравнения yчн следует искать в виде константы.
Учитывая, что первая и вторая производная от константы равны нулю, при подстановке в уравнение (3.26) yчн =const получим уравнение (3.34)
![]() (3.34)
(3.34)
![]() (3.35)
(3.35)
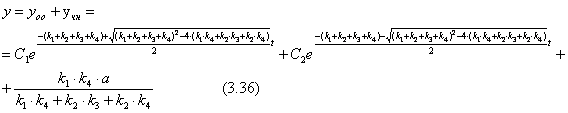
Для нахождения х необходимо выразить ![]() из выражения (3.36).
из выражения (3.36).
![]() (3.37)
(3.37)
Подставим найденное значение y и ![]() в выражение (3.17) и выразим x:
в выражение (3.17) и выразим x:
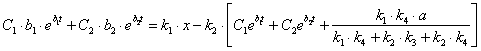 (3.38)
(3.38)
![]()
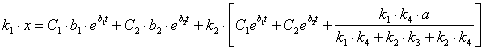 (3.38)
(3.38)
![]() (3.39)
(3.39)
![]()
 (3.40)
(3.40)
Подставим найденные значения y и z в выражение (1) и выразим x:

Нахождение С1 и С2.
Учитывая, что ![]() , после подстановки в выражение (3.36) получим
, после подстановки в выражение (3.36) получим
выражение (3.37).
![]() (3.42)
(3.42)
Учитывая, что ![]() , после подстановки в выражение (3.40) получим:
, после подстановки в выражение (3.40) получим:
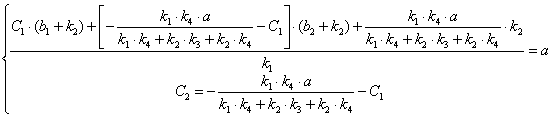 (3.43)
(3.43)
![]()
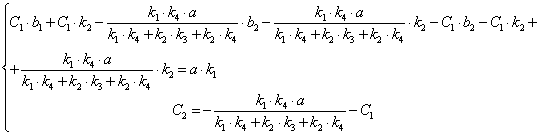 (3.44)
(3.44)
![]()
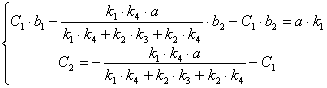 (3.45)
(3.45)
![]()
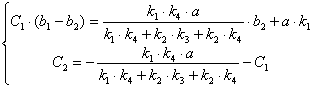 (3.46)
(3.46)
![]()
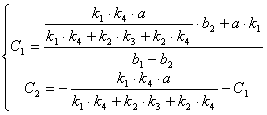 (3.47)
(3.47)
![]()
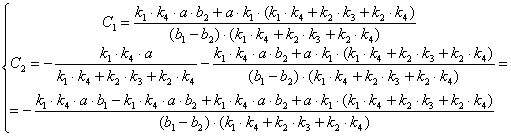 (3.48)
(3.48)
![]()
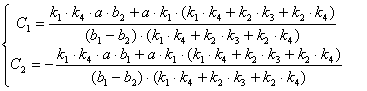 (3.49)
(3.49)
Подставим найденные С1 и С2 в уравнения (3.40):
 (3.50)
(3.50)
Подставим найденные С1 и С2 в уравнения (3.13):
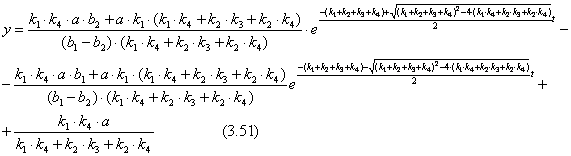
Для выражения z подставим найденные значения для С1 и С2 в уравнение (3.41) и получим уравнение (3.52).

Выразим k2, k3, k4 через k1. Константа скорости образования комплекса ДНК.Pr (k1) является функцией от T, свойствами реагирующих веществ и других параметров (и) определяется (опытным путем) для каждой реакции индивидуально. Так как при каждом ПЦР цикле концентрация ПЦР-продукта увеличивается в 1,8 – 1,95 раза, то мы можем задать k1=0,05, при котором 85% ДНК будет превращается в комплекс ДНК.Pr.
Константа скорости разрушения комплекса ДНК.Pr (![]() ) может быть вычислена с помощью константы равновесия (K) из выражения (3.53).
) может быть вычислена с помощью константы равновесия (K) из выражения (3.53). ![]() , (3.53)
, (3.53)
где K – константа равновесия.
Константа равновесия может быть рассчитана из уравнения (3.54).
![]() , (3.55)
, (3.55)
где ![]() – изменение энергии Гибса в результате реакции.
– изменение энергии Гибса в результате реакции.
Разность энергий Гибса до и после реакции может быть выражена согласно уравнения (3.33).
![]() , (3.56)
, (3.56)
где ![]() H – изменение энтальпии до и после реакции,
H – изменение энтальпии до и после реакции, ![]() S - изменение энтропии до и после реакции.
S - изменение энтропии до и после реакции.
Энтальпия и энтропия могут быть вычислено по принципу адитивности [51] из которого следует уравнение 3.57 и 3.58.
![]() (3.57)
(3.57)
![]() (3.58)
(3.58)
Выразим ![]() из выражений (3.31-3.35):
из выражений (3.31-3.35):
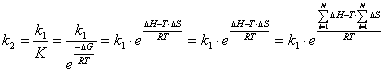 (3.59)
(3.59)
Константа скорости образования комплекса ДНК.Pr* (k3) зависит от:
- вероятности столкновения двух молекул (ДНК и Pr);
- энергетического барьера;
- энергии молекул;
- (пространственных затруднений).
Так как компоненты реакций образования специфических и неспецифических комплексов ДНК.Pr одинаковы (ДНК и Pr), то вероятность столкновения ДНК и Pr, энергетический барьер, энергия молекул и константа скорости одинаковы. Тоесть ![]() .
.
Константа скорости разрушения комплекса ДНК.Pr* (![]() ) может быть выражена через
) может быть выражена через ![]() по тому же алгоритму, как и
по тому же алгоритму, как и ![]() , которое выражалось через
, которое выражалось через ![]() в уравнении (3.59):
в уравнении (3.59):
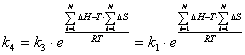 (3.60)
(3.60)
Теперь, когда мы выразили x, y, z через ![]() ,
, ![]() ,
, ![]() ,
, ![]() , a,
, a, ![]() и выразили
и выразили ![]() ,
, ![]() ,
, ![]() через
через ![]() , мы можем построить график изменения концентрации одноцепочечной ДНК (x = f1(
, мы можем построить график изменения концентрации одноцепочечной ДНК (x = f1(![]() )), комплекса ДНК-праймер (y = f2(
)), комплекса ДНК-праймер (y = f2(![]() )), не специфического комплекса ДНК-праймер z = f3(
)), не специфического комплекса ДНК-праймер z = f3(![]() ) со временем. Значение энтальпии и энтрапии мы брали из [78]
) со временем. Значение энтальпии и энтрапии мы брали из [78] ![]() ,
, ![]() ,
, ![]() . Данный график представлен на рисунке № 3.1, расчеты проводились при температуре 60ºС. Для построения графика была использована программа Mathcard 2001 Professional.
. Данный график представлен на рисунке № 3.1, расчеты проводились при температуре 60ºС. Для построения графика была использована программа Mathcard 2001 Professional.
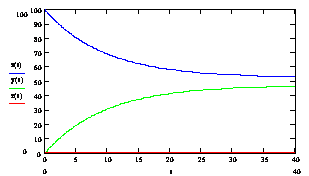
Рис № 3.1. Зависимость концентрации ssДНК и комплексов ДНК.Pr, ДНК.Pr* от времени, при 60 ºС.
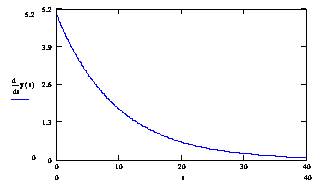
Рисунок № 3.2. График изменения скорости накопления комплекса ДНК-праймер (![]() ) во времени при температуре 60ºС.
) во времени при температуре 60ºС.
Из графиков 3.1. и 3.2. видно, что отжиг праймеров при температуре 60 С0 (близкой к температуре плавления) имеет преимущества и недостатки. К преимуществам можно отнести то, что отжиг праймеров идет с высокой специфичностью. Концентрация комплексов неспецифического комплекса ДНК- праймер (ДНК.Pr*) очень низкая С(ДНК.Pr*)t=40c = 0.23 %. К недостаткам можно отнести то, что скорость прироста концентрации комплексов ДНК.Pr быстро падает (см. рис. 3.2). Поэтому нам необходимо изменять температуру, поддерживая оптимальную скорость образования комплексов ДНК.Pr на том уровне, который обеспечит нужный уровень специфичности.
Получим математическую модель процесса отжига праймеров. При условии, что температура является функцией от времени, то k2 и k4 также являются функциями от времени. Описывать данную систему будет система дифференциальных уравнений:
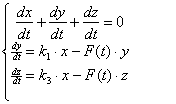 (3.61)
(3.61)
Данное уравнение проблематично решить аналитически, его можно решить для конкретно заданной функции, например такой как ![]() или
или ![]() , где
, где ![]() - температура,
- температура, ![]() - время. Однако решение данного уравнения будет чрезвычайно громоздким. Поэтому гораздо проще задаться ступенчато изменяющейся температурой и при переходе на новую температуру переносить предыдущие значения концентрации ssДНК, комплексов ДНК.Pr и ДНК.Pr*. Для этого мы можем воспользоваться формулами (3.36, 3.40, 3.41) и задаться новыми краевыми условиями y(0) = y0 и x(0) = x0.
- время. Однако решение данного уравнения будет чрезвычайно громоздким. Поэтому гораздо проще задаться ступенчато изменяющейся температурой и при переходе на новую температуру переносить предыдущие значения концентрации ssДНК, комплексов ДНК.Pr и ДНК.Pr*. Для этого мы можем воспользоваться формулами (3.36, 3.40, 3.41) и задаться новыми краевыми условиями y(0) = y0 и x(0) = x0.
После подстановки y(0) = y0 в выражение (3.36) получим уравнение (3.62).
![]() (3.62)
(3.62)
После подстановки x(0) = x0 в выражение (3.40) получим:
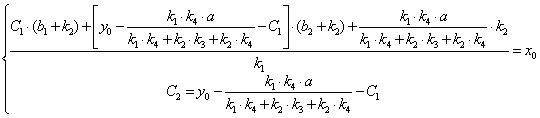 (3.63)
(3.63)
![]()
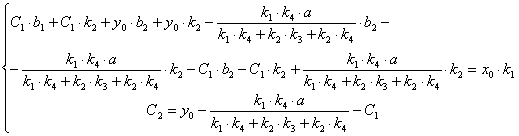 (3.64)
(3.64)
![]()
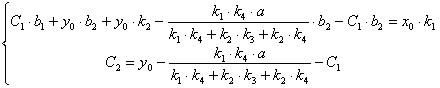 (3.65)
(3.65)
![]()
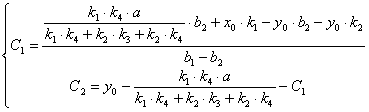 (3.66)
(3.66)
![]()
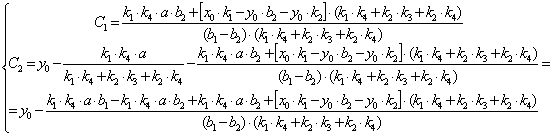 (3.67)
(3.67)
![]()
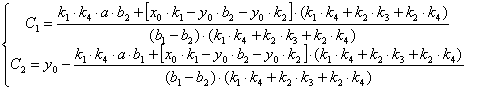 (3.68)
(3.68)
Подставим найденные значения С1 и С2 в уравнения (3.36, 3.40, 3.41) и выразим x (уравнение(3.69)), y(уравнение(3.70)), z(уравнение(3.71)).
 (3.69)
(3.69)


Теперь мы можем построить график № 3.3 зависимости концентрации ssДНК и комплексов ДНК.Pr, ДНК.Pr* от времени, при ступенчато изменяющейся температуре 10с при 600С, 30с при 580С. Для построения графика используем программу Mathcard 2001 Professional. Для сравнения приведем график зависимости концентрации ssДНК и комплексов ДНК.Pr, ДНК.Pr* от времени, при температуре 600С (рис. 3.1) и 580С (рис. 3.5).
Если сравнить рисунки № 3.7, 3.9 , то видно, что отжиг праймеров гораздо выгоднее проводить при повышающейся температуре от 580С до 600С. так как отношение концентраций ДНК.Pr к ДНК.Pr* составит 230, в то время как при понижении температуры от 600С до 580С отношение концентраций ДНК.Pr к ДНК.Pr* составит 80.
Как видно из графика на рисунке 3.3, снижение температуры от 600С до 580С приводит к увеличению скорости накопления комплекса ДНК.Pr, но в тоже время ведет к стремительному увеличению концентрации неспецифического комплекса ДНК.Pr*. Необходимо отметить, что согласно рисунка № 3.5, скорость накопления специфических и неспецифических комплексов ДНК-праймер меняется скачкообразно в сторону увеличения.

Рис 3.3. Зависимость концентрации ssДНК и комплексов ДНК.Pr, ДНК.Pr* от времени, при изменяющейся температуре 10с при 600С, 30с при 580С.
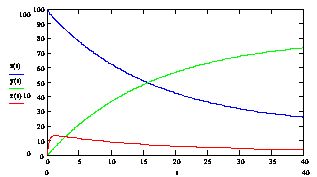
Рис 3.4. Зависимость концентрации ssДНК и комплексов ДНК.Pr, ДНК.Pr* от времени, при 580С.
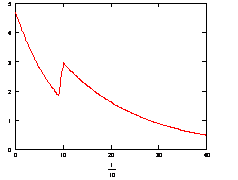
Рис. 3.5. График изменения скорости накопления комплекса ДНК-праймер (![]() ) во времени при изменяющейся температуре 10с при 600С, 30с при 580С.
) во времени при изменяющейся температуре 10с при 600С, 30с при 580С.
Рассмотрим процесс отжига праймеров при возрастающей температуре. На рисунке 3.7. представлен график зависимости концентрации комплексов ДНК.Pr к ДНК.Pr*, от времени, при увеличивающейся температуре 10с при 580С, 30с при 600С. Как видно на рисунке 3.7. повышение температуры с 580С до 60ºС приводит к резкому снижению концентрации неспецифических комплексов ДНК-праймер, что, безусловно, имеет положительный эффект, но этот положительный эффект затмевается резким снижением скорости прироста специфических комплексов ДНК-праймер рисунок 3.7. Если не повышать температуру, то скачкообразного снижения скорости прироста специфических комплексов ДНК-праймер наблюдаться не будет (рис 3.8.)
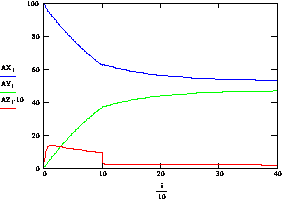
Рис. 3.6. Зависимость концентрации комплексов ДНК.Pr к ДНК.Pr*, от времени, при изменяющейся температуре 10с при 58ºС, 30с при 60ºС.
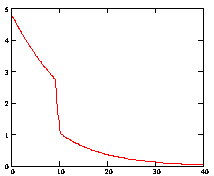
Рис. 3.7. Зависимость отношения концентрации комплексов ДНК.Pr к ДНК.Pr*, от времени, при изменяющейся температуре 10с при 580С, 30с при 600С.
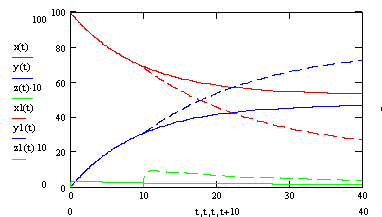
Рис. 3.8. Зависимость концентрации ssДНК и комплексов ДНК.Pr, ДНК.Pr* от времени, при изменяющейся температуре 10с при 600С, 30с при 580С. Пунктирной линией показано изменение концентрации ssДНК и комплексов ДНК.Pr, ДНК.Pr* от времени, при температуре 580С.
Перед нами была поставлена цель увеличения выхода продукта и специфичности. Для максимизации выхода продукта необходимо постепенно снижать температуру отжига праймеров тем самым поддерживать скорость накопления специфического продукта, но снижать ее такими темпами, чтобы не допускать снижения специфичности ниже 99%.
Рассмотрим процесс отжига праймеров при ступенчато возрастающей температуре. На рис 3.9. представлен график зависимости концентрации ssДНК и комплексов ДНК.Pr, ДНК.Pr* от времени, при изменяющейся температуре 5с при 60,5ºС, 15с при 58ºС и 20с при 56ºС. Проведение многоступенчатого отжига праймеров с постепенным снижением температуры позволяет поддерживать высокую скорость образования специфических комплексов ДНК-праймер (рисунок 3.10), при высокой специфичности (рисунок 3.11).
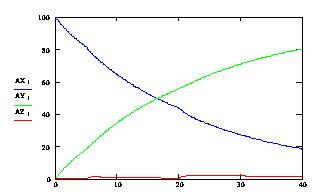
Рис. 3.9. Зависимость изменения концентрации ssDNA (синяя линия), специфического комплекса ДНК.Pr (зеленая линия), неспецифического комплекса ДНК.Pr* (красная линия) от времени, при изменяющейся температуре 5с при 60,5ºС, 15с при 58ºС и 20с при 56ºС.
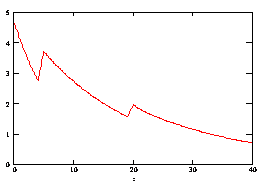
Рисунок 3.10. Скорость изменения концентрации комплекса ДНК-праймер в течении времени.
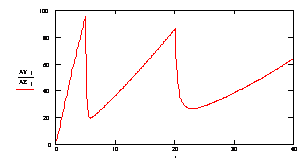
Рисунок 3.11. Изменение отношения концентрации специфического комплекса ДНК-праймер к неспецифическому комплексу ДНК-праймер в течении времени.
Приведенная модель моделирует процесс отжига праймеров и рассматривает 4 процесса:
1) ssДНК+Pr![]() ssДНК.Pr (3.72)
ssДНК.Pr (3.72)
2) ssДНК.Pr![]() ssДНК+Pr (3.73)
ssДНК+Pr (3.73)
3) ssДНК+Pr![]() ssДНК*.Pr (3.74)
ssДНК*.Pr (3.74)
4) ssДНК*.Pr![]() ssДНК+Pr (3.75)
ssДНК+Pr (3.75)
Но модель не рассматривает процесс элонгации цепи и ДНК:
5) ssДНК.Pr + dNTP![]() ДНК + H3PO4 (3.76)
ДНК + H3PO4 (3.76)
6) ssДНК*.Pr + dNTP ![]() ДНК* + H3PO4 (3.77)
ДНК* + H3PO4 (3.77)
7) ssДНК + ssДНК ![]() ДНК (3.78)
ДНК (3.78)
Последняя реакция (3.78) идет гораздо медленнее остальных реакций, так как С(ssДНК) << C(Pr), поэтому мы не будем ее рассматривать. Скорость образования ПЦР – продукта лимитируется концентрацией ssДНК.Pr и фермента. Скорость образования продукта полимеразной цепной реакции описывается уравнением (3.79).
V5=k5.C(ssДНК.Pr) . C(E) (3.79)
где: C(E) – концентрация фермента.
Так как С(E) не изменяется во времени, то мы можем внести ее в k5 и получить уравнение (3.80)
V5=k5.C(ssДНК.Pr) (3.80)
Зависимость скорости синтеза от температуры представлена в таблице № 1.11. Исходя из данных таблицы 1.11., мы вывели корреляционное уравнение (3.81) , которое будет использовано для интерполяции.
k5= 0.02![]() (3.81)
(3.81)
Исходя из вышеперечисленного, построим математическую модель данного процесса. Данный процесс будет описываться системой диф. уравнений (3.82).
 (3.82)
(3.82)
Для простоты записи заменим, что x = С(ДНК), y = C(ДНК.Pr), z = C(ДНК*.Pr), A = C(ДНК), B=C(ДНК*), a = C(ДНК)0 и получим:
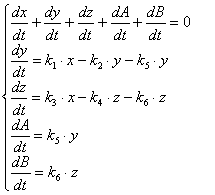 (3.83)
(3.83)
В данном уравнении k2, k4, k5, k6 - функции от температуры, а температура является функцией от времени. Аналитическое решение данного уравнения весьма проблематично, поэтому применим итерационный метод. Суть данного метода заключается в том, что мы сначала задаем начальные параметры x=100, ![]() , а затем пересчитываем значения x, y, z, a, b при
, а затем пересчитываем значения x, y, z, a, b при ![]() c согласно системе уравнений (3.84).
c согласно системе уравнений (3.84).
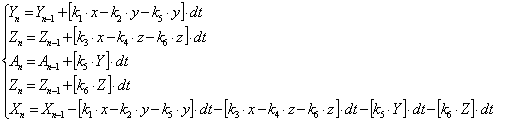 (3.84)
(3.84)
После ![]() таких шагов мы получаем значения концентраций, отвечающие 40с отжига праймеров. Расчеты проводятся при помощи программы GraphBuilder, которая была разработана нами в среде программирования Borland Delphi 7.0. Данный программный продукт интегрирован с базой данных Genome на базе MS SQL Server при помощи технологии ActiveX Data Objects, в которой хранятся данные о состоянии системы через каждые 100 млс, что эквивалентно десяти тысячам вычислений. Эти данные используются для построения графиков.
таких шагов мы получаем значения концентраций, отвечающие 40с отжига праймеров. Расчеты проводятся при помощи программы GraphBuilder, которая была разработана нами в среде программирования Borland Delphi 7.0. Данный программный продукт интегрирован с базой данных Genome на базе MS SQL Server при помощи технологии ActiveX Data Objects, в которой хранятся данные о состоянии системы через каждые 100 млс, что эквивалентно десяти тысячам вычислений. Эти данные используются для построения графиков.
Данный метод имеет преимущества и недостатки. К преимуществам можно отнести то, что мы можем задавать температуру как функцию от времени и других параметров любой степени сложности. К недостаткам можно отнести меньшую точность, чем при аналитическом решении системы дифференциальных уравнений, но это нивелируется за счет низкого значения dt, которое обеспечивает высокую точность.
Проанализируем динамику изменения концентрации компонентов ПЦР системы во время амплификации при помощи программы GraphBuilder при различных функциональных зависимостях температуры от времени. Результаты анализа представлены в Табл. № 3.2.
Таблица № 3.2.
| № |
Начальная температура |
Конечная температура |
Функция |
Концентрации реагентов |
||||
| X |
Y |
Z |
A |
B |
||||
| 1 |
55 |
55 |
|
14,4 |
48,2 |
2,1 |
31,2 |
4,1 |
| 3 |
55 |
60 |
|
29.2 |
34.4 |
0.1 |
34.6 |
1.7 |
| 2 |
60 |
55 |
|
16.4 |
47.2 |
2 |
33,6 |
0,9 |
| 4 |
60 |
60 |
|
41.6 |
28.6 |
0.1 |
29.5 |
0.2 |
| 5 |
58 |
58 |
|
21,9 |
41,9 |
0,3 |
35,2 |
0,65 |
Если учесть что при элонгации цепи ДНК.Pr и ДНК.Pr* полностью превратятся в соответствующий ПЦР – продукт, то необходимо рассматривать не A и B, а A+Y и B+Z. Результаты представлены в таблице № 3.3.
Таблица № 3.3.
| № |
Функция |
Концентрации реагентов |
A+Y / B+Z |
Рис. |
||
| X |
A+Y |
B+Z |
||||
| 1 |
|
14,4 |
79,4 |
6,18 |
12,9 |
3.12 |
| 2 |
|
29.2 |
69,0 |
1,79 |
38,6 |
3.13 |
| 3 |
|
16,4 |
80,83 |
2,81 |
28,74 |
3.14 |
| 4 |
|
41.6 |
58,02 |
0,35 |
163,62 |
3.15 |
| 5 |
|
21,9 |
77,13 |
0,97 |
79,22 |
- |
Из первой и четвертой строки таблицы 3.3 видно, что с ростом температуры увеличивается специфичность и уменьшается выход продукта, что согласуется с практикой и литературными данными. Наша задача подобрать такой температурный режим, при котором специфичность была бы выше 99%, а выход продукта максимальный. При этом целесообразно рассматривать не только линейное изменение температуры (общее уравнение можно записать в виде (3.85)), но и другие функции:
- экспоненциальное изменение температуры (общее уравнение можно записать в виде (3.86));
- степенное изменение температуры (3.87);
- логарифмическое изменение температуры (3.89).
![]() (3.85)
(3.85)
![]() (3.86)
(3.86)
![]() (3.87)
(3.87)
![]() (3.88)
(3.88)
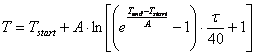 (3.89)
(3.89)
При поиске функции оптимального отжига праймеров будем изменять:
- температуру старта отжига праймеров с 55 до 65 ºС с шагом 1 градус;
- конечную температуру отжига праймеров, которую проба достигнет через 40 секунд с 55 до 65 ºС с шагом 1 градус;
- коэффициент A, который определяет вид функции изменения температуры, для каждой функции меняется по разному.
Таким образом мы смоделировали процесс отжига праймеров при изменяющейся температуре согласно функций (3.86 – 3.89). Интервалы варьирования и шаг варьирования температуры старта, температуры окончания отжига праймеров и кофицента А, а также количество проанализированных функций, представлены в таблице 3.4.
Таблица 3.4.
| Вид функции |
Интервал варьирования температуры старта ºС |
Шаг варьирования температуры старта ºС |
Интервал варьирования температуры окончания ºС |
Шаг варьирования температуры окончания ºС |
Интервал варьирования коэффициента А |
Шаг варьирования коэффициента А |
Количество проанализированных функций |
| (3.86) |
[55; 65] |
1 |
[55; 65] |
1 |
[-3; 3] |
0.1 |
11.11.60 = 7260 |
| (3.87) |
[55; 65] |
1 |
[55; 65] |
1 |
[0; 3] |
0.1 |
11.11.30 = 3630 |
| (3.88) |
[55; 65] |
1 |
[55; 65] |
1 |
(0; 3] |
0.1 |
11.11.30 = 3630 |
| (3.89) |
[57; 67] |
0.2 |
[57; 67] |
0.2 |
[-2; -0.45] |
0.05 |
50.50.31 =77500 |
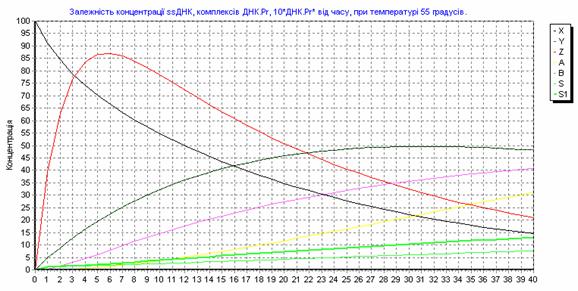
Рис. 3.12. Зависимость концентрации ssДНК и комплексов ДНК.Pr, ДНК.Pr* от времени, при 550С.
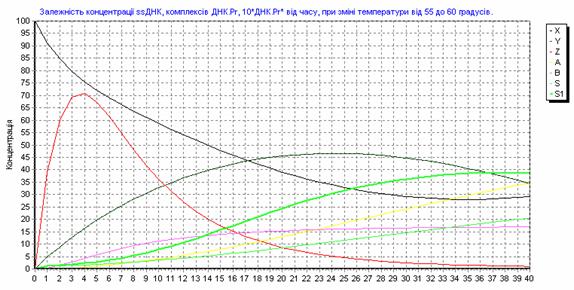
Рис. 3.13. Зависимость концентрации ssДНК и комплексов ДНК.Pr, ДНК.Pr* от времени, при изменении температуры от 55 до 600С.
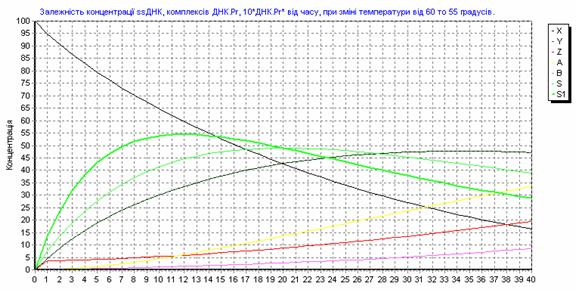
Рис. 3.14. Зависимость концентрации ssДНК и комплексов ДНК.Pr, ДНК.Pr* от времени, при изменении температуры от 60 до 550С.
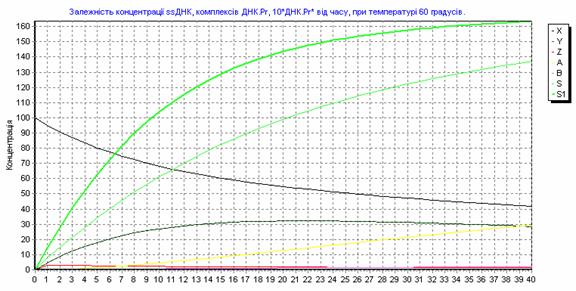
Рис. 3.15. Зависимость концентрации ssДНК и комплексов ДНК.Pr, ДНК.Pr* от времени, при 600С.
Результаты расчетов концентрации реагентов записывались с шагом 1 секунда в базу данных. Так как мы записывали количественные показатели состояния системы через каждую секунду, то получили 40 записей на один вариант изменения температуры амплификации и 3680800 записей на 92020 вариантов изменения температуры амплификации.
Скрипт, представленный в приложении 2, позволил выбрать те записи, где специфичность превысила 99% и выход продукта превысил 75%. Результат выполнения скрипта представлен в таблице 3.5.
Таблица 3.5.
| Results |
||||||||||||||
| Acceleration |
T |
X |
Y |
Z |
A |
B |
S |
TC |
AY |
BZ |
S1 |
Process |
Tsta |
Tend |
| -2 |
40 |
24,09 |
41,19 |
0,35 |
33,96 |
0,41 |
83,11 |
58 |
75,15 |
0,76 |
99,01 |
(3.86) |
60 |
58 |
| -1,9 |
40 |
24,17 |
41,15 |
0,35 |
33,92 |
0,41 |
83,64 |
58 |
75,07 |
0,76 |
99,15 |
(3.86) |
60 |
58 |
| -4 |
40 |
23,66 |
41,67 |
0,34 |
33,91 |
0,41 |
82,1 |
58 |
75,58 |
0,76 |
99,73 |
(3.86) |
61 |
58 |
| -3,9 |
40 |
23,72 |
41,65 |
0,35 |
33,87 |
0,41 |
82,59 |
58 |
75,52 |
0,76 |
99,91 |
(3.86) |
61 |
58 |
| -3,8 |
40 |
23,79 |
41,62 |
0,35 |
33,83 |
0,41 |
83,09 |
58 |
75,46 |
0,75 |
100,09 |
(3.86) |
61 |
58 |
| -3,7 |
40 |
23,86 |
41,6 |
0,35 |
33,79 |
0,4 |
83,59 |
58 |
75,39 |
0,75 |
100,26 |
(3.86) |
61 |
58 |
| -3,6 |
40 |
23,93 |
41,57 |
0,35 |
33,75 |
0,4 |
84,11 |
58 |
75,32 |
0,75 |
100,43 |
(3.86) |
61 |
58 |
| -3,5 |
40 |
24,01 |
41,54 |
0,35 |
33,7 |
0,4 |
84,65 |
58 |
75,24 |
0,75 |
100,6 |
(3.86) |
61 |
58 |
| -3,4 |
40 |
24,09 |
41,51 |
0,35 |
33,65 |
0,4 |
85,19 |
58 |
75,16 |
0,75 |
100,77 |
(3.86) |
61 |
58 |
| -3,3 |
40 |
24,17 |
41,48 |
0,35 |
33,6 |
0,39 |
85,75 |
58 |
75,08 |
0,74 |
100,92 |
(3.86) |
61 |
58 |
| -0,4 |
40 |
24,233 |
40,559 |
0,304 |
34,458 |
0,446 |
77,286 |
58,2 |
75,017 |
0,75 |
100,001 |
(3.89) |
59,6 |
58,2 |
| -0,8 |
40 |
24,222 |
41,162 |
0,352 |
33,866 |
0,397 |
85,266 |
58 |
75,028 |
0,749 |
100,14 |
(3.89) |
61,4 |
58 |
| -0,8 |
40 |
24,243 |
41,164 |
0,352 |
33,845 |
0,396 |
85,512 |
58 |
75,008 |
0,748 |
100,259 |
(3.89) |
61,6 |
58 |
| -0,75 |
40 |
24,124 |
41,211 |
0,351 |
33,913 |
0,4 |
84,689 |
58 |
75,124 |
0,751 |
100,014 |
(3.89) |
63 |
58 |
| -0,75 |
40 |
24,127 |
41,211 |
0,351 |
33,911 |
0,4 |
84,715 |
58 |
75,122 |
0,751 |
100,027 |
(3.89) |
63,2 |
58 |
| -0,75 |
40 |
24,128 |
41,212 |
0,351 |
33,909 |
0,4 |
84,735 |
58 |
75,121 |
0,751 |
100,037 |
(3.89) |
63,4 |
58 |
| -0,75 |
40 |
24,13 |
41,212 |
0,351 |
33,907 |
0,4 |
84,75 |
58 |
75,119 |
0,751 |
100,045 |
(3.89) |
63,6 |
58 |
| -0,75 |
40 |
24,131 |
41,212 |
0,351 |
33,906 |
0,4 |
84,762 |
58 |
75,118 |
0,751 |
100,051 |
(3.89) |
63,8 |
58 |
| -0,75 |
40 |
24,132 |
41,212 |
0,351 |
33,905 |
0,4 |
84,771 |
58 |
75,118 |
0,751 |
100,055 |
(3.89) |
64 |
58 |
| -0,75 |
40 |
24,132 |
41,212 |
0,351 |
33,905 |
0,4 |
84,778 |
58 |
75,117 |
0,751 |
100,059 |
(3.89) |
64,2 |
58 |
| -0,75 |
40 |
24,133 |
41,212 |
0,351 |
33,904 |
0,4 |
84,783 |
58 |
75,117 |
0,751 |
100,061 |
(3.89) |
64,4 |
58 |
| -0,75 |
40 |
24,133 |
41,213 |
0,351 |
33,904 |
0,4 |
84,787 |
58 |
75,116 |
0,751 |
100,063 |
(3.89) |
64,6 |
58 |
| -0,75 |
40 |
24,133 |
41,213 |
0,351 |
33,903 |
0,4 |
84,79 |
58 |
75,116 |
0,751 |
100,065 |
(3.89) |
64,8 |
58 |
| -0,75 |
40 |
24,133 |
41,213 |
0,351 |
33,903 |
0,4 |
84,792 |
58 |
75,116 |
0,751 |
100,066 |
(3.89) |
65 |
58 |
| -0,75 |
40 |
24,134 |
41,213 |
0,351 |
33,903 |
0,4 |
84,794 |
58 |
75,116 |
0,751 |
100,067 |
(3.89) |
65,2 |
58 |
| -0,75 |
40 |
24,134 |
41,213 |
0,351 |
33,903 |
0,4 |
84,796 |
58 |
75,116 |
0,751 |
100,068 |
(3.89) |
65,4 |
58 |
| -0,75 |
40 |
24,134 |
41,213 |
0,351 |
33,903 |
0,4 |
84,797 |
58 |
75,115 |
0,751 |
100,068 |
(3.89) |
65,6 |
58 |
| -0,75 |
40 |
24,134 |
41,213 |
0,351 |
33,903 |
0,4 |
84,798 |
58 |
75,115 |
0,751 |
100,069 |
(3.89) |
65,8 |
58 |
| -0,75 |
40 |
24,134 |
41,213 |
0,351 |
33,903 |
0,4 |
84,798 |
58 |
75,115 |
0,751 |
100,069 |
(3.89) |
66 |
58 |
| -0,75 |
40 |
24,134 |
41,213 |
0,351 |
33,903 |
0,4 |
84,799 |
58 |
75,115 |
0,751 |
100,069 |
(3.89) |
66,2 |
58 |
| -0,75 |
40 |
24,134 |
41,213 |
0,351 |
33,902 |
0,4 |
84,799 |
58 |
75,115 |
0,751 |
100,07 |
(3.89) |
66,4 |
58 |
| -0,75 |
40 |
24,134 |
41,213 |
0,351 |
33,902 |
0,4 |
84,799 |
58 |
75,115 |
0,751 |
100,07 |
(3.89) |
66,6 |
58 |
| -0,75 |
40 |
24,134 |
41,213 |
0,351 |
33,902 |
0,4 |
84,8 |
58 |
75,115 |
0,751 |
100,07 |
(3.89) |
66,8 |
58 |
| -0,75 |
40 |
24,134 |
41,213 |
0,351 |
33,902 |
0,4 |
84,8 |
58 |
75,115 |
0,751 |
100,07 |
(3.89) |
67 |
58 |
Из таблицы видно, что наибольший интерес представляют функции (3.86) и (3.89). В функции (3.86) температура отжига праймеров от времени находится в экспоненциальной зависимости. График зависимости температуры от времени для функции (3.90) представлен на рисунке (3.12).
В функции (3.89) температура отжига праймеров изменяется в логарифмической зависимости, график зависимости температуры от времени для функции (3.91) представлен на рисунке (3.13).
![]() (3.90)
(3.90)
![]() (3.91)
(3.91)
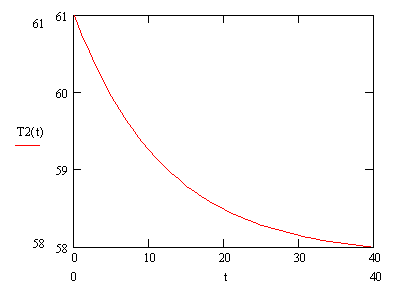
Рисунок 3.16.
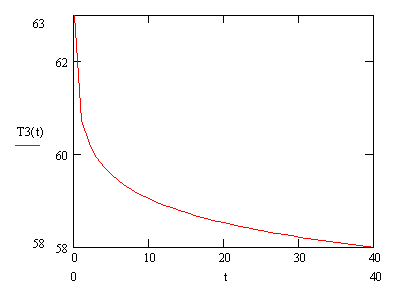
Рисунок 3.17.
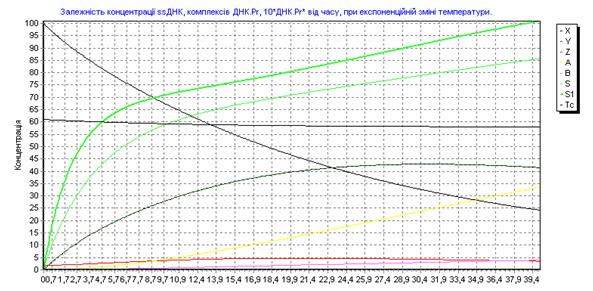
Рис. 3.18. Зависимость концентрации ssДНК и комплексов ДНК.Pr, ДНК.Pr* от времени, при изменении температуры по формуле (3.90).
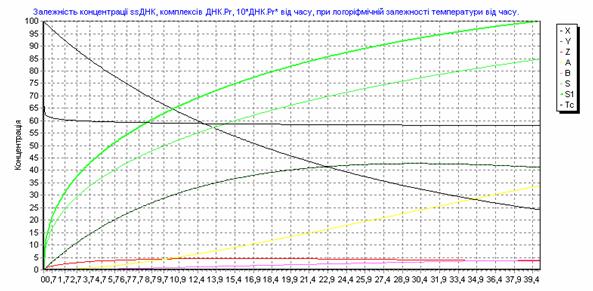
Рис. 3.19. Зависимость концентрации ssДНК и комплексов ДНК.Pr, ДНК.Pr* от времени, при изменении температуры по формуле (3.91).
4. ВЫВОДЫ.
1. Был созданный банк ДНК больных на ФКУ и членов их семей из разных регионов Украины.
2. Освоены методы: выдиления ДНК, амплификации, рестрикции, вертикального и горизонтального электрофореза.
3. Подобранные условия для амплификации in vitro и рестрикции последовностельности седьмого экзона гена ФАГ.
4. Получена маниматическая модель отжига праймеров и оптимальный температурный режим амплификации.
5. Изучение алельного полиморфизма VNTR-локуса 3'-нетрансльо-ваної области гена ФАГ показало довольно высокую инфрмативность полиморфизма для проведения внутрисемейного анализа и возможность его использования в качестве маркера для проведения ДНК-диагностики фенилкетонурии. 33 % проанализированных семей оказались информативными, 33 % полуинформативными, 33 % неинформативными.
